
Tardigrad
| Tardigrad | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

tardigrad Milgiene tardigradum | ||||||||||||
| sistematik | ||||||||||||
| ||||||||||||
| Bilimsel ad | ||||||||||||
| Tardigrada | ||||||||||||
| Spallanzani , 1777 | ||||||||||||
| sınıflar | ||||||||||||
Su ayıları (Tardigrada) - olarak da bilinir , su ayılar - bir formu hayvan suşu içinde deri değiştirme hayvanlar ( Ecdysozoa). Genellikle bir milimetreden küçük olan sekiz bacaklı hayvanlar, görünüşleri ve Almanca konuşulan ülkelerde adlarına yol açan beceriksiz hareket tarzları nedeniyle bir şekilde ayıları andırıyor . Bilimsel adı (bir arada Latin Tarduşlar 'yavaş' ve gradus 'adım' ) geri yavaş harekete geçer.
Dünya çapında denizde, tatlı suda veya karada nemli habitatlarda yaşarlar; özellikle orada yosun yastıklarında yaygındırlar . Hayvanların bir özelliği, aşırı çevresel koşullarda hayatta kalabilecekleri ölüm benzeri bir durum olan kriptobiyozdur . Tardigradlar, bitki hücrelerinin içeriğiyle beslenebilir ve ayrıca diktikleri ve emdikleri yuvarlak solucanlar (Nematoda) veya rotiferler (Rotifera) gibi küçük hayvanlar üzerinde yırtıcıdır . Tardigrad genellikle cinsel olarak ürer . Bununla birlikte, bazı türler de partenogenetik olarak , yani erkeklerin katılımı olmadan çoğalır ; bu durumda dişilerin yumurtaları döllenmeden gelişir.
Tardigradların bir sonraki yakın akrabaları muhtemelen Panarthropoda taksonunu oluşturdukları uzuvlar (Arthropoda) ve kadife solucanlardır (Onychophora) .
yapı

Tardigrade büyüklüğü bazı yavru 50 mikrometre ve 1.5 mm arasındadır türleri cinsi Macrobiotus böylece hayvanlar edilen, ancak genellikle yaklaşık olarak 100 ila 500 mikrometre olan sayılır şekilde meiofauna . Nispeten dolgun, silindirik şekilli bir gövdeye sahiptirler, karınları yassıdır ve yüzeyde her biri bir çift bacak ve bir kafa parçasına sahip dört vücut bölümünden oluşuyormuş gibi görünür. Bununla birlikte, derinin en dış tabakasındaki enine kıvrımların neden olduğu bu dış izlenim yanıltıcıdır: Vücudun geri kalanından yalnızca hafifçe ayrılmış ve farklılaşmış olan kafa, muhtemelen bir değil, üç iç içe geçmiş bölümden oluşur; beden ise dört değil, beş parçadan oluşmalıdır.
Deniz türleri çoğunlukla renksiz veya göze çarpmayan beyazdan griye renkliyken, kara ve tatlı su formları genellikle kırmızı, sarı, yeşil, pembe, mor veya siyah gibi renklere sahiptir. Renk, dış derideki, kütiküldeki pigmentlerden veya vücut boşluğunun, hemokoelomanın veya bağırsağın renkli içeriğinden kaynaklanır . Boyalar da sıklıkla bağırsaktan hemokoeloma göç eder ve oradan kütikülde birikir.
Karakteristik olarak, tardigrad dokuların çoğu çirkindir , yani oluşturdukları hücre sayısı genetik olarak belirlenir. Boyut olarak büyüme, bu nedenle, hücre sayısındaki bir artışla değil, yalnızca tek tek hücrelerin kendi büyümeleriyle gerçekleşebilir. İlk hücre bölünmesinden sonra iki yavru hücreden biri (deneysel müdahale ile) öldürülürse, anatomik olarak eksiksiz ve yaşayabilir bir hayvan gelişecektir, bu daha sonra normalin sadece yarısı kadar hücre içerir.
ekstremiteler

Dört gövde segmentinde, karnın sol ve sağ kenarında (ventrolateral olarak), bir "güdük bacak" ortaya çıkar, aynı zamanda ilgili güdük ayak grubunda da olduğu gibi, gövdenin eklemsiz bir çıkıntısı . Bazen, daha sonra koksa ("kalça"), femur ("uyluk"), tibia ("shin") veya tarsus ("ayak") olarak adlandırılan bacağın ayrı bölgeleri arasında bir ayrım yapılır . Bacaklar sadece biraz farklı olduğundan, bu terimlerin anatomik temeli sorgulanabilir; her halükarda, ancak bunlar tamamen tanımlayıcı olarak anlaşılmalıdır, yani eklembacaklıların karşılık gelen bacak bileşenleri ile evrimsel bir ilişki göstermezler.
Birçok deniz türü, bacaklarını teleskopla birbirine geçirebilir; Tüm tardigradların bacağının içinde çalışan kısa kaslar bu konuda onlara yardımcı olur. Bacağın sonunda genellikle dört ila sekiz, daha nadiren on üç geri çekilebilir pençe vardır, bunlar bazen ince bacak çıkıntılarına, "ayak parmaklarına" oturur ve özel bezler, "pençe bezleri" tarafından salgılanır . Bazı türlerde pençeler yapışkan disklere ikincildir; Orada hiçbir pençeleri içinde cinsi Apodibius . Genellikle dördüncü çift bacağın pençeleri, ilk üç çiftinkiyle zıt yönü gösterir.
deri

Vücut, hücresel olmayan bir dış katman, kütikül ve tek tek hücrelerden oluşan altta yatan bir iç katman olan epidermis tarafından dışarıya sınırlandırılmıştır . Tardigrad deride kamçılı hücreler görülmez.
Kütikül 0.5 mikrometre kalınlığındadır ve üç katman, dış oluşur epicuticle 0.1 ila yaklaşık mikrometre altta yatan bir kalınlığı olan intracuticle yaklaşık 0.2 mikrometre arasında bir kalınlığa ve bir son ile procuticle 0 .25 mikron ile ilgilidir. Bu oluşan kitin , çeşitli glikoproteinler , polisakkaritler , mukopolisakkaritler , lipid ve lipoprotein ve aynı zamanda pek çok karasal ve tatlı su türleri de pigmentler içerir. Genellikle tümsekler, dikenler veya çukurlarla süslenmiştir ve bazı türlerde sırtta (dorsal), daha nadiren yanlarda (yanal) sertleştirilir , çok nadiren ayrıca kalın zırh plakaları, skleritler oluşturmak için karın (ventral) üzerinde de bulunur. ; bunlar daha sonra eklembacaklılarda olduğu gibi eklemlenir. Kütikül, bacak pençeleri ve ön ve arka bağırsağın astarı ile birlikte, eski stilettolarda bulunan ağız parçaları da dahil olmak üzere düzenli olarak derilenir .
Epidermis bir Eutelian'dan oluşur, yani her zaman aynı sayıda hücreden oluşan bir tür içinde, üstündeki kütikül tarafından salgılanan basit hücre tabakası. Vücut boşluğundan sadece bazal lamina adı verilen ince bir hücre dışı ayırıcı tabaka ile ayrılır.
hemokoelom
Vücut boşluğu gerçek bir sölom değildir , yani embriyonik mezoderme kadar izlenebilen bir doku ile kaplı değildir . Tüm organlar ondan sadece ince bazal lamina ile ayrılır. Vücut boşluğu nedenle olarak ifade edilir psödomonasa bir şekilde, “kan” dolu olduğu için, ya da hemocoeloma . Gerçek bir sölom sadece tek gonad (gonad) çevresinde bulunabilir.
Kan renksizdir ve oksijen taşımaya hizmet etmez. Vücut hareketleriyle dolaşır; ancak ayrı kan damarları veya kalp gibi gerçek bir kan dolaşımı yoktur. Bu amaçla, 200'e kadar hücre ile doldurulur, hemositler serbest yüzer veya bazal laminaya sabitlenir.İlk görevleri besinlerin depolanması gibi görünmektedir; bakterilerle mücadelede de rol oynayabilirler.
kas sistemi
Yakından ilişkili kolombdan farklı olarak, tardigraddaki kaslar asla kalın kas katmanlarında görünmez. Bunun yerine, gövde ve uzuvların kasları, kütiküle bağlı izole, ince uzunlamasına kaslardan oluşur, bunlar sadece birkaç, hatta bazen tek bir kas hücresinden oluşur. Ya bir hidro -iskelet gibi sıvı dolu bir balon gibi davranan hemocoel'e karşı ya da insan kolunun fleksör ve ekstansör kasları gibi birbirine karşı antagonist olarak çalışırlar . Tardigradda halka kasları bulunmaz. Kas hücreleri çoğunlukla düzdür, nadiren çizgilidir, ikincisi muhtemelen orijinal durumdur; Yaygın bir hipoteze göre, düz kaslar birçok türde ancak ikincil olarak kırsal yaşama uyum olarak gelişmiştir.
nefes almak
Kaslar çalışmaları için oksijene ihtiyaç duyarlar , bunun tardigrad tarafından ortalama tüketiminin saatte 0,05 ile 0,1 milimetre küp ve vücut ağırlığının miligramı arasında olduğu tahmin edilmektedir. Bununla birlikte, vücut yüzeyinin ve hacminin çok uygun oranı nedeniyle, ayrı solunum organları gerekli değildir ve buna göre mevcut değildir. Tüm gaz değişimi deriden basit difüzyonla gerçekleşebilir, ancak bunun ince bir su tabakasıyla kaplanması gerekir.
Sindirim organları
Sindirim sistemi vücudun önünden arkasına doğru uzanan ve ağız boşluğu, boğaz (yutak), yemek borusu, orta bağırsak ve rektum olarak ayrılabilen uzun bir tüpten oluşur. İlk üç bölüm, ön bağırsak (yemek borusu) olarak gruplandırılmıştır, çünkü kütikül astarı, kütikül arka bağırsak astarı ile birlikte, her tüy dökümü ile dökülür.
Etçil veya omnivor türlerde (etobur ve omnivor), ağız açıklığı genellikle vücudun ön ucunun (terminal) ucunda, bitki veya organik atıklarla beslenen türlerde (otçul ve detritivor) ise, genellikle ön ucun biraz gerisindedir Ventral taraf (ventral-subterminal). Genellikle 10 ila 30 sert, küboid çıkıntılı, lamel adı verilen bir halka ile çevrilidir ve bazen iç içe geçebilen bir ağız konisi üzerine oturur.
Bir tükürük bezi, sol ve sağdaki bitişik tübüler ağız boşluğuna açılır, bu sadece sindirim salgılarını salgılamakla kalmaz, aynı zamanda tardigradın özelliği olan stilettoları da sentezler. Bunlar sert, çoğunlukla düz, ancak bazen de genellikle ilgili tükürük bezinin iç kısmında (lümen) bulunan, ancak keskin uçları ile ağız boşluğuna çıkabilen kavisli iğnelerdir. Kalınlaşmış arka uçlarına bağlanan ön ve ekartör kasları, onları germek veya geri çekmek için kullanılabilir. Enine stiletto tutucular onları ağız boşluğuna bağlarken ön uçları geri çekildiğinde kendi stiletto kılıflarında durur. Etçil türlerde, genellikle otçul formlardan biraz daha belirgindir.
Farinks , vücut boşluğundan bir bazal lamina ile ayrılan, radyal olarak dışa doğru uzanan çizgili epitel kas hücrelerinden oluşur . Boğazın iç kısmı, lümen ise bir kütikül ile kaplıdır ve Y-şekilli, triadidik kesiti sayesinde sıvıyı emmek için kullanılan verimli negatif basınç üretimi için idealdir. bağırsaklara yiyecek. Bu pompa prensibi, rotiferler gibi yakından ilişkili olmayan diğer hayvanlarda da bağımsız olarak gelişmiştir .
Muhtemelen mukus salgılayan kübik hücrelerden oluşan kısa bir yemek borusu , bazen yüzey alanını arttırdığına inanılan yanal çıkıntılara (mikrovilli) sahip tek bir epitel dokusu tabakasından oluşan orta bağırsağa bağlanır . Bağırsaklara girerken, gıda genellikle işlevi hassas bağırsak dokusunu yabancı cisimlerin zarar görmesinden korumak olan bir peritrofik zar ile sarılır . Midgut hidrolitik enzimler salgılar , besinlerden besinleri emer, yağ veya polisakkarit şeklinde depolar ve muhtemelen atık maddelerin yok edilmesinde de görev yapar. Pek çok türde , muhtemelen kommensal olarak , yani ev sahibini etkilemeden yaşayan , ancak bazı durumlarda aynı zamanda bir besin kaynağı olarak da hizmet eden geniş bir bakteri florası vardır.
Yiyecek kalıntıları, bir kütikül ile kaplı kısa bir rektumda toplanır, muhtemelen yeniden değiştirilir ve daha sonra orta hattaki (medial-ventral) son bacak çiftinin biraz önünde, karın üzerindeki anüs yoluyla dış dünyaya salınır . . Birçok türde, yalnızca aşağıda daha ayrıntılı olarak açıklanan Malpigh bezleri değil , aynı zamanda dişilerin fallop tüpleri de rektuma açılır ; Bu durumda bağırsağın son bölümüne kloak denir.
boşaltım organları
Sindirilemeyen kalıntılar, metabolik atık ürünler veya kirleticiler çeşitli şekillerde dış dünyaya salınabilir. Bir yandan, birçok kirletici epidermis tarafından üstteki kütikül içine yerleşir ve daha sonra bir sonraki tüy dökümü sırasında onunla birlikte dökülür . Ek olarak, tükürük bezleri görünüşte tüy dökümü sırasında bir boşaltım işlevi üstlenir. Bağırsak sadece besinleri emmekle kalmaz, aynı zamanda kirleticileri vücut boşluğundan bağırsağın iç kısmı olan bağırsak lümenine bırakır.
Eutardigrada sınıfına atanan birçok türün ayrıca Malpigh bezleri olarak bilinen üç, çok nadiren dört özel organı vardır . İkisi yan tarafta (yan), biri bağırsağın arkasında (dorsal) bulunur. Orta ve rektumun birleştiği yerde açılırlar ve büyük olasılıkla azotlu atıkları ortadan kaldırmak için kullanılırlar. Yapısal olarak aynı adı taşıyan böcek bezlerine benzemelerine rağmen, muhtemelen homolog organlar değillerdir , yani evrimsel anlamda ortak bir öncül yapıya geri dönmezler. İkinci büyük sınıf olan heterotardigrada'nın orijinal olarak kabul edilen deniz türlerinde bulunmadığından, tatlı su ve karadaki yaşama evrimsel bir adaptasyon olarak kabul edilirler.
Heterotardigrada'nın bazı kırsal formları, Malpigh bezleri yerine, ikinci ve üçüncü çift bacakların tabanında açılan ve muhtemelen bir boşaltım işlevine sahip olan, karın üzerinde yer alan özel organlara sahiptir. Gerçek nefridia , oldukça gelişmiş boşaltım organları ise tardigradda oluşmaz.
Gergin sistem
Tardigradın sinir sistemi, kafada ön bağırsağın etrafında dönen bir sinir halkasından ve göbek tarafında geriye doğru uzanan iki çift sinir kordonundan oluşur ve her gövde segmentinde çapraz bağlantılar yoluyla ip merdiven sinir sistemi denilen bir sinir sistemi oluşturur .
Ağız üst kısmında uzanan bir supraözofageal gangliyon ve aynı subözofagealin altında bulunan bir ön sinir halkası , sindirim kanalının yanından geçen iki sinir hücresi kümesi sinir bantları bir halka oluşturacak şekilde birbirine bağlanır ve ilkel bir " beyin düşünülebilir". Üst faringeal ganglion, arkada bulunan ve arka uca (kaudal olarak) bakan iki çift beyin lobundan, bir iç ve bir dış çiftin yanı sıra stilettoların yanında bulunan bir çift beyin lobundan oluşur. Dış arka çift, varsa, gözleri innerve eder ve ayrıca kafa, sirri ve klavada bulunan diğer duyu organlarını sinirlerle besler. Aynı zamanda alışılmadık bir şekilde birinci gövde segmentinin ganglionlarına da bağlıdır . Bununla birlikte, ilgili eklembacaklılarda olduğu gibi, bunlar öncelikle geniş sinir bağlarıyla yeraltı gangliyonuna bağlanır.
Tardigrad beyin, en az üç segmentin kaynaşmasından başın kökeninin bir göstergesi olarak yorumlanabilen, orijinal olarak bağımsız birkaç ganglionun kaynaşmasından ortaya çıkar. Yakın akraba eklembacaklıların beyni ayrıca, proto-, deuto- ve tritoserebrum olmak üzere üç beyin bölgesini oluşturan birkaç gangliyondan oluşur. Bu nedenle bazen karşılık gelen yapıların homolojiler , yani tardigrad ve eklembacaklıların ortak atalarına giden dokular olduğu varsayılır . Daha yakın tarihli elektron mikroskobik çalışmalar bu görüşle çelişir ve bunun yerine tardigradın tüm beyninin evrimsel terimlerle eklembacaklıların ön beyin kısmına tekabül ettiği sonucuna varır.
Karın üzerinde çalışan iki sinir kordonu gövde sinir sistemini oluşturur. Yeraltı gangliyonunda ortaya çıkarlar ve hayvanın arka ucuna paralel olarak uzanırlar. Her bir sinir kordonu, dört bacak taşıyan segmente ve belki de başka bir bacaksız genital segmente karşılık gelen dört veya beş gangliona sahiptir. Bir bacak segmentinin iki gangliyonu, enine sinir bağlantıları ile birbirine bağlanır.
Duyu organları
Hepsi olmasa da çoğu tardigratın kırmızı veya siyah nokta şeklinde gözleri vardır. Pigment kupası oocelli olarak tasarlanmıştır, yani her bir göz, içbükey tarafı vücut yüzeyine ve dolayısıyla ışığa bakan iki fotoreseptör hücre ile çevrili fincan şeklinde bir pigment hücresinden oluşur. Beynin dış, arka lobları tarafından sinirlerle beslenirler ve özel formlarında sadece tardigratta bulunurlar.
Ek olarak, bazı türlerin gövde bölümlerinde muhtemelen kimyasal veya dokunma uyaranlarına tepki veren kıl şeklinde sensillalar vardır . İplik şeklindeki sensillalar, cirri , özellikle birçok deniz türünde başın arkasında bulunur ve muhtemelen dokunma organlarıdır, oysa klava , biraz daha kalın ve içi boş iplikler, muhtemelen kemoreseptörlerdir . Birçok türde , muhtemelen çevrenin algılanmasında da bir işlevi olan ağız açıklığı, papilla çevresinde siğil şeklinde yükselmeler vardır .
Üreme organları
Tardigrade, temel olarak, sindirim sisteminin üzerinde yer alan ve bağlarla arka vücut duvarının ön ucuna bağlanan gerçek sölomik doku ile çevrili sadece bir eşleşmemiş gonada sahiptir.
Flagellalı sperm , erkeklerin testislerinde oluşur. Ondan çıkan iki sperm iletkeni, karın üzerinde cinsel açıklıkta birleşen gonopore, vücudun orta hattında anüsün önünde yer alır, genellikle tüp benzeri çıkıntı yapar ve dış dünyaya açılır. Sperm merdiveninin işlevsel olarak açıklanamayan kopyalanması, bir gonadın evrimsel kaybının bir göstergesi olarak görülüyor.
Dişi yumurtalık ise, türe bağlı olarak bağırsağın sağında veya solunda bulunan yalnızca bir fallop tüpüne sahiptir. Bir sınıfın türü olan heterotardigrada'da ağzı, genellikle anüsün önünde bulunan ayrı bir gonopore içinde bulunurken, diğer sınıfın türü olan eutardigrada'da ise arka bağırsakta bulunur ve böylece bir kloak haline gelir. . Birçok dişide, çiftleşme sırasında erkeklerin spermlerini alan ve yumurtlayana kadar depolayan bir veya iki seminal vezikül vardır.
Hermafroditik bireyler, hem sperm hem de yumurta hücrelerinin olgunlaştığı ve paylaşılan bir yumurta veya seminal kanal yoluyla salınabilen ovotestis olarak bilinen bir gonada sahiptir .
dağıtım
Tardigradlar, Antarktika dahil tüm kıtalarda ve tüm okyanuslarda yaşar . Orta Avrupa yağmur oluklarında ve ayrıca düzenli olarak buzlu kutup havuzlarında veya tropikal yağmur ormanlarında , Himalayalar'da 6000 metreden fazla yükseklikte , Güney Sandwich Adaları gibi uzak adalarda , 4690 metre derinliğindeki abisal bölgede bulunabilirler. Hint Okyanusu'nun tabanında veya Atlantik'in ortasında sürüklenen kahverengi algler üzerinde . Tüm iklim kuşaklarında görülmekle birlikte, ana yayılış alanı kutup ve ılıman enlemlerdedir.
Bir Biyocoğrafi yapı olabilir ancak ilişkilendirilebilir familya ve cins düzeyinde kabul edilmesi , eski kıta ayrılması ile Pangea'nın içine Gondwana'dan ve Laurasia . Toplam on cins ve 22 tür kozmopolittir, yani tüm dünyada bulunur. Triyas'ın jeolojik çağından önce var olan bir Pangea faunasının kalıntıları olarak kabul edilirler . Diğer türlerin çoğu, mekansal olarak sınırlı bir menzile sahiptir.
yetişme ortamı

Tardigrad, habitatlarına göre temel olarak deniz ( deniz ), tatlı su ( limnic ) veya karada yaşayan ( karasal ) olarak tanımlanabilir, bu nedenle son iki kategori arasındaki ayrım sadece belirsizdir, bu nedenle toplu limnotkarasal terimi sıklıkla kullanılır. . Tüm tardigradlar, bazıları dehidrasyona karşı oldukça dirençli olmasına rağmen, aktif bir yaşam için ince bir su tabakasına bağlıdır.
Deniz habitatı
Deniz türleri hem tuzlu hem de acı suda yaşar ve gelgitler arası bölgeden sığ su bölgesinden okyanusların dipsiz düzlüklerine kadar bulunabilir; en az bir tür, Mariana Çukuru'nun zemininde hakim olan su basıncına dayanabilir . Deniz dibi çamurluysa, oradaki bentik tardigradlar genellikle çevrelerine, kısaltılmış uçlara sahip solucan biçimli güçlü bir gövdeyle adapte olmuşlardır; Öte yandan, yarık ve yarıklara sahip kumlu veya çakıllı alt toprakta, iyi gelişmiş kütük bacakları olan hayvanların bulunma olasılığı daha yüksektir. Bir tür görünüşte uzman olan manganez nodüller , bir yaşam alanı olarak ise alglerin geniş halı veya hayvanlar da, diğerleri taş mercan (Scleractinia), Bryozoa , midye (Bivalvia), woodlice (Isopoda), kıskaç (Cirripedia), deniz kestaneler (Echinoidea) veya Deniz kameraları (Holothuroidea) kısmen ortak , yani konakçıyı etkilemeden, kısmen de parazitik olarak yaşar . Sargasso Denizi'ndeki açık okyanusun ortasında yüzen Sargassum alglerinde en az bir tür yaşıyor . Karaya doğru, hayvanların bireysel kum taneleri arasında yaşadığı hemen hemen tüm plajlarda toprağın üst santimetrelerinde belirgin bir kum boşluğu faunası vardır. Birkaç tür varsa, bunlar çoğunlukla birbirinden nem veya sıcaklık farklılıklarıyla ayrılabilen farklı mikro habitatlara dağılır. Karasal formlara geçiş, deniz likenlerinde yaşayan, genellikle yüksek gelgitte ulaşılan ve normalde sadece tuzlu sprey ile ulaşılan su seviyesinin üzerindeki kayalar üzerine yerleşmiş tardigradlar tarafından oluşturulur .
tatlı su habitatı

Kelimenin tam anlamıyla limnik türler, göllerde, göletlerde, göletlerde ve hatta bireysel su birikintilerinde olduğu kadar akan sularda da yaşarlar. Tardigradlar genellikle bir göl içindeki dipte yaşayan kum aralığı faunasının bir parçasını oluşturur; 150 metreye kadar derinlikte buluntular bilinmektedir. Hayvanlar çoğunlukla toprağın en üst, oksijen içeren santimetrelerinde yaşarlar, kendilerini gevşek kumun hayvanlara ciddi şekilde zarar verebileceği, ancak çoğunlukla biraz daha derinde göl kıyısına doğru bulurlar. Ek olarak, algler ve su bitkileri kolonizedir ve bazen göl yüzeyindeki nilüferlerde tardigradlar bulunabilir .
Genellikle bireysel bireyler yağmur oluklarında izlenebilir; hayvanlar muhtemelen çatıdaki yosunlarla yıkanır, böylece karasal olarak kabul edilirler. Sonuçta, kaplıcalar ayrıca bazı türlerin yaşadığı bir tatlı su habitatı oluşturur.
Habitat arazisi

En önemli karasal habitatlar yosun çimleridir; orada yaşayan türler yosun seven veya briyofilik olarak bilinir . Ancak yosunlar suyu aralıklarda depoladığından, aynı zamanda su habitatları olarak da görülebilirler. Yosunların farklı bölgelerinde genellikle farklı türler bulunur; yosunların toprakta kök saldığı köksap tabakasının tardigrad faunası ve örneğin fotosentetik olarak aktif, ancak dehidrasyona eğilimli dış tabaka aynı değildir. Ayrıca likenlerde veya bromeliadlar (Bromeliaceae), ibrik bitkileri (Nepenthaceae) gibi uygun çiçekli bitkilerde saksafon ( Saxifraga ) Mannsschild ( Androsace ) veya taraklamada ( Dipsacus ) bulunan hayvanlar . Bu bitkilerin tümünde su, yaprak köklerinde veya bitkinin çanak şeklindeki diğer kısımlarında birikir; tardigrade için minyatür bir yaşam alanı haline gelirler.
Tardigradlar ayrıca genellikle ormanların yaprak döküntülerinde veya zeminin kendisinde bulunur ve kayın ormanlarının özellikle popüler olduğu görülmektedir. Biraz daha alışılmadık bir habitat , yüksek dağlardaki buzullardır : orada, karanlık toz birikintileri veya ince taneli molozlar gün boyunca ısı emilimine ve dolayısıyla yüzeyin geçici olarak çözülmesine neden olabilir; tardigrad , elde edilen sulu kriyokonit tabakasında da bulunabilir. Kentsel habitatlar bugüne kadar pek çalışılmamıştır.
Tardigradlar, yalnızca kendi substratlarında ince bir su filmiyle kaplandıklarında aktiftirler, böylece çöller gibi kurak alanlarda yaşayamazlar. Bununla birlikte, birçok karasal tür, kuraklığa son derece dayanıklıdır ve bu nedenle düzenli kurak dönemlere ve aşırı sıcaklıklara dayanabilir. Bazı rotiferlerle (Rotifera) paylaştıkları bu temel ekolojik avantaj, çevredeki nem içeriğinin en azından kısmen büyük tercih olan güçlü dalgalanmalara maruz kaldığı yuvarlak solucanlar (Nematoda) gibi rakip hayvanlar üzerinde en iyi şekilde oynayabilirler. çok kısa sürede kuruyan veya suyla dolup taşabilen yosunlar ve likenler açıklanmıştır. Diğer birkaç hayvanın erişebildiği bu habitatların kolonizasyonu, tardigratın büyük evrimsel başarısının önemli bir nedeni olarak kabul edilir.
Nüfus yoğunlukları
Popülasyon yoğunlukları hakkında kesin bilgiler sadece birkaç tür için mevcuttur ve şimdiye kadar sadece örneklerde mevcuttur: kumlu plajlarda santimetre küp kum başına 35'e kadar birey vardı, topraklarda santimetre kare başına 30'a kadar birey sayıldı, yosunlar ise yosun yastıklar başına yaklaşık 200 kişi Beklendiği gibi, santimetre kare çok yoğun nüfuslu bir habitatı temsil ediyor. Ancak kural olarak, nüfus yoğunlukları bu maksimum değerin oldukça altındadır. Şimdiye kadarki yetersiz veri durumuna dayanarak söylenebileceği kadarıyla, deniz türlerinin bireysel sayıları genellikle kara veya tatlı su formlarından çok daha küçüktür.
Nüfus yoğunlukları sıcaklık ve nemden, besin kaynağından, parazit ve yırtıcıların sayısından veya çevresel kirleticilerden etkilenebilir ve limnotkarasal türlerde mevsime bağlı olarak çoğunlukla dalgalanır, ilk zirve ilkbahar ve yaz başında ve ikinci bir zirve ile mevsime bağlı olarak değişir. sonbaharda zirve.
Ulaşım ve hareket
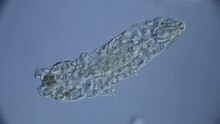
Tardigrade, aktif hareket yoluyla yalnızca çok alt düzeyde yayılır; çoğu hayvan bunun yerine rüzgar, su veya hayvanlar tarafından yeni potansiyel habitatlara getirilir. Bu pasif taşıma, öncelikle hayvanların yumurtalarını, ayrıca kistleri ve fıçıları etkiler - çevresel aşırılıklardan büyük ölçüde veya aşırı derecede bağımsız olan yaşam aşamaları.
Deniz türleri okyanus akıntılarında sürüklenebilir, bu da muhtemelen dış derilerinde yelken benzeri özel çıkıntılarla onlara yardımcı olur. Limnotrestrial türler bazen taşan akarsular veya erimiş su ile aynı şekilde taşınır. Geçici kuru habitatlar, yumurtaların veya kistlerin rüzgarla ( anemochory ) yayılmasına izin verirken, daha küçük alanlar su sıçramalarında köprülenebilir. Grönland açıklarında bir fırtına sırasında, muhtemelen fırtına tarafından önceden fırlatılan düşen yağmur damlalarında yetişkin hayvanlar bile tespit edilebilir. Tahminen, yumurtaların veya kistlerin yapıştığı böcekler veya kuşlar da tardigradları yeni habitatlara ( zochories ) getirir .
Aktif hareket için tüm türler ince bir su tabakasına ihtiyaç duyar. Daha sonra, ilgili columbus'tan farklı olarak, yalnızca çiftler halinde değil, örneğin kum taneleri üzerinde gezinmek veya yosun yastıklarına tırmanmak için bireysel olarak da hareket ettirilebilen ilk üç gövde bölümünün bacaklarını kullanırlar. Bacaklardaki pençeler veya yapışkan diskler, ilgili alt tabakayı kavramak için kullanılır. İlk üç çift bacaktan farklı olarak, iki arka uzuv yere tutunmaya ve bazı türlerde geriye doğru hareket etmeye hizmet eder; farklı pençe hizalaması yoluyla bu değiştirilmiş işleve uyarlanırlar. Deneysel olarak ölçülen en yüksek koşu hızı saatte 17,7 santimetredir; değeri, Latin türetilen bir grup, Tardigrada, Latin adı yapan Tarduşlar 'yavaş' ve Gradus anlamına gelen 'yavaş strider', uygun görünen 'adım',.
Fototaksi , yani ışık kaynaklarına gidiş ve dönüş hareketi henüz yeterince araştırılmamıştır. Genç hayvanlar fotokinetik olarak olumsuz tepki veriyor gibi görünüyor , yani ışık ışımasına daha hızlı hareketlerle ve kendiliğinden yön değişiklikleriyle tepki veriyorlar, belirli bir kaçınma tepkisi tespit edilmeden. Işık radyasyonu genellikle termal radyasyon ve ardından suyun buharlaşması ile ilişkili olduğundan, bu davranış belki de genç hayvanlar için daha tehdit edici olan dehidrasyon riski ile ilgilidir.
Bir deniz türü aktif olarak yüzebilir; kütikülü çan şeklindedir ve büzülme yoluyla yönlendirilmiş bir su jetini dışarı atabilir; hayvanlar geri tepme ilkesine göre denizanası gibi hareket ederler .
beslenme

Tardigradların çoğu , vahşi veya likenlerde buldukları alg hücreleri başta olmak üzere vejeteryan bir diyetle beslenir . Yosunların bitki hücreleri ise, orada kalıcı olarak yaşayan hayvanlarda bile nadiren besin spektrumuna aittir. Alglerin yanı sıra toprakta yaşayan türler , içerdiği bakteri ve mantar sporları ile birlikte organik atıkları da yutar ; bununla birlikte, bazı formlar da tamamen veya kısmen yırtıcı olarak yaşar. Av çeşitleri arasında protozoa , rotiferler (Rotifera) ve yuvarlak solucanlar (Nematoda) ve ayrıca diğer tardigradlar bulunur. Bazı deniz türleri deniz cevizlerinin veya midyelerin kabuğunda ektoparazitler olarak yaşarlar . Karada, tardigradlar, sürekli dehidrasyon riski nedeniyle parazitik bir yaşam biçimine zayıf bir şekilde adapte olurlar; Kara salyangozlarında endoparazit olabilen burada sadece bir tür bilinmektedir .
Tardigrade, yemek için ağız konilerini etkilenen bitki hücresine, avlarının derisine veya ev sahibinin vücut duvarına bastırır. İğne keskinliğinde stileler ilerletilerek bunlar daha sonra delinir veya delinir ve tüm hücre veya vücut içeriği emilir. Özellikle büyük türler ise sadece sıvı gıda almakla kalmaz, aynı zamanda avlarını bir bütün olarak emer; bu özellikle daha küçük tekerlekli hayvanları ve tardigradları etkiler.
Yırtıcı hayvanlar, parazitler ve ortaklar
Tardigrade'nin başlıca yırtıcıları, diğer tardigrade türleri, rotiferler (Rotatoria) ve nematodların yanı sıra akarlar (Acari), örümcekler (Araneae), yay kuyrukları (Collembola), çeşitli böcek larvalarıdır , ayrıca solucanlar (Lumbricidae) gibi spesifik olmayan "otlayan" organizmalardır. veya çeşitli tatlı su yengeçleri.
Bazı tardigradlar , ince hücre iplikleri ( hyphae ) avlarının yakalandığı ve daha sonra yakalanan hayvanlara dönüşen halkalar halinde iç içe geçen etçil mantarların kurbanı olur . İpliksi mantar cinsi Atricordyceps'in aseksüel aşaması olan Harposporium gibi diğer mantarlar , muhtemelen tardigradlar tarafından yenildikten sonra bağırsakta filizlenen ve avlarını içeriden sindiren aseksüel sporlar olan conidia verir. Mantar ayrıca hayvanların kütiküllerine dışarıdan da nüfuz edebilir. Tardigradın diğer önemli mantar parazitleri, lazımlık mantarı Sorochytrium milnesiophthora ve boyunduruk mantarları Ballocephala sphaerospora ve Ballocephala verrucospora'dır .
Protozoalar arasında muhtemelen birkaç parazit vardır ; bu henüz ayrıntılı olarak çalışılmamıştır, ancak tercihen tardigradları enfekte eden Pyxidium tardigradum siliatları . Muhtemelen bir symphoriont olarak yaşıyor , bu da yalnızca konakçısı tarafından yayılabileceği, ancak kendisi için tehlikeli görünmediği anlamına geliyor.
Birçok simbiyotik bakteri genellikle deniz tardigradında bulunur ; Karada yaşayan formlar da çoğunlukla zengin bir bakteriyel bağırsak florasına sahiptir. Proteobacterium Xanthomonas campestris , bitki önemli bir zararlı, tardigrade tarafından iletilen inanılmaktadır.
deri değiştirme
Tardigrade, yalnızca düzenli deri değiştirme yoluyla boyut olarak büyüyebilir . Stilelerle birlikte hücresel olmayan dış deri (kütikül), ön ve arka bağırsağın astarı ve bacak pençeleri dökülür. Eski stilettolar ve ağız boşluğu ve ön bağırsağın kütikulasının dışarı atılması gerçeğiyle, tüy dökümü her zaman hayvanların ön ucunda başlar. Vücut geçici olarak kasıldığında, eski deriden ayrılır ve daha sonra soyulur ve boş bir gömlek ( exuvium ) olarak geride bırakılır. Genellikle 5 ila 10 gün süren bu süreçte hayvanlar, herhangi bir yiyecek alamayacakları stilesiz simpleks aşamasındadır . Tüy dökümü sırasında, alttaki epidermisten yeni kütiküller oluşurken, tükürük bezlerinde yeni stilettolar sentezlenir ve pençeler özel pençe bezleri tarafından oluşturulur.
Bireysel bireylerde yaşamları boyunca 13'e kadar tüy dökümü tespit edilebilir; Yaşam boyu büyümeyi mümkün kılmalarının yanı sıra, iç vücut basıncını düşürmeye, kütikülde depolanan atık maddeleri uzaklaştırmaya veya parazitleri gidermeye de hizmet ederler. Birçok tür, yumurtalarını tüy döken gömleklere bırakır.
Direnç aşamaları
Tardigradların çoğu, kuraklık, soğuk dönemler, suyun tuzluluğundaki keskin dalgalanmalar veya oksijen eksikliği dönemlerinde hayatta kalabilmek için benzersiz adaptasyonlar geliştirmiştir. Sağlamlığa bir katkı onların Eutelia'sıdır: Embriyonik gelişimden sonra, hassas fazları ile hücre bölünmesi neredeyse hiç gerçekleşmez ( germ hattı dışında ). Bu zaten çok yüksek bir radyasyon direncine neden olur ( sterillik dışında ), karşılaştırma için → Radyasyon hasarı bölümündeki tabloya bakın . Ayrıca bazı türler morfolojik değişikliklerle mevsimlere uyum sağlarken bazıları kalın duvarlı kistler oluşturabilir. Bununla birlikte, aşırı adaptasyon şekli , hayvanların daha fazla metabolik aktivitenin kaydedilemeyeceği ölüme yakın bir duruma geçtiği kriptobiyoz olarak adlandırılır . Direncin tüm aşamaları, olumsuz çevresel koşullardan kurtulmaya hizmet eder ve böylece tardigrade'e diğer hayvan grupları üzerinde evrimsel bir rekabet avantajı sağlar. Ayrıca hayvanların yeni potansiyel habitatlara pasif taşınmasında da rol oynarlar. Tardigrad aynı zamanda uzayda hayatta kalabildiği bilinen ilk hayvandır.
siklomorfoz
Siklomorfoz, çevresel koşullardaki döngüsel değişikliklere yanıt olarak vücut şeklindeki düzenli değişikliktir. Morf olarak bilinen hayvanların yaşam evrelerinin çoğu mevsime bağlı olarak birinden diğerine değişir. Şimdiye kadar, siklomorfozis sadece cinslerden deniz su ayıları bilinir Halobiotus , Amphibolus ve Hypsibius . Şimdiye kadar incelenen en iyi örnek , Grönland'ın gelgitler arası bölgesinde yaşayan Halobiotus crispae türünde bulunur : Burada, pseudosimplex adı verilen bir kış morfu , bir yaz morfundan ayırt edilebilir. İlki donmaya yakın sıcaklıklara dayanabilir ve düşük sıcaklıklarda bile hareketli kalır, ancak sterildir; sadece çok daha az dirençli olan yaz morfu verimlidir.
kistler
Özellikle tatlı suda yaşayan, aynı zamanda yosunları veya yaprak döküntülerini kolonize eden türler, kist olarak bilinen direnç aşamaları geliştirebilir. Bunu yapmak için hayvanlar önceki vücut boyutlarının yüzde 20 ila 50'sine kadar küçülür, metabolizmalarını azaltır ve bazı durumlarda iç organlarını da parçalar. Bu sürece, sonunda hayvanın kütikül katmanlarından oluşan çok duvarlı bir kaplama ile çevrelendiği üç taneye kadar tamamlanmamış, hemen ardışık tüy dökümü eşlik eder. Bu durumda hayvanlar bir yıldan fazla yaşayabilir. Çevre koşulları değişir değişmez, hayvanlar 6 ila 48 saat içinde kendilerini yeniden kafeslerinden kurtarabilirler.
Temel olarak kistler sadece suda oluşur; aşağıda belirtilen tamburlar kadar dayanıklı değildirler ve bunların aksine su içeriğinden dolayı ısıya dayanıklı da değildirler.
anoksibiyoz
Anoksibiyoz, oksijen eksikliğine toleransı tanımlamak için kullanılan terimdir. Bu solunum gazının konsantrasyonu çok düşükse, ozmoregülasyon bozulur , hayvanlar su yutar ve önemli ölçüde şişer. Tüm metabolizma artık anaerobik olarak gerçekleşmelidir, toksik atık ürünler vücut boşluğunda birikir. Tardigradların çoğu hala böyle bir durumda 3 ila 5 gün hayatta kalabilir; Echiniscoides cinsinden bireysel bireylerin , çürüyen midyelerden oluşan kapalı bir test tüpünde 6 aya kadar hayatta kalabildikleri bile bildirilmiştir . Normal duruma dönüş, anoksibiyotik aşamanın süresine bağlı olarak birkaç dakika ile birkaç saat arasında sürer.
Anoksibiyoz, örneğin büyük statik su kütlelerinde, oksijen konsantrasyonunun geçici olarak keskin bir şekilde düşebileceği habitatlarda özellikle önemlidir. Gelgitler arası bölgede alg halıları üzerinde yaşayan Tardigrade, gelgitin alçaldığı zamanlarda aşırı oksijen eksikliğine de maruz kalır ve evrimsel anlamda, belki de bu adaptasyonu ilk elde edenler arasında yer alır.
ozmobiyoz
Osmobiosis, suyun tuzluluğundaki dalgalanmaları tolere etme yeteneğidir. Tardigrad, çoğu hayvan gibi, ancak sudaki iyon konsantrasyonu belirli sınırlar içindeyse yaşayabilir . Bununla birlikte, özellikle gelgit bölgesinde yaşayan türler, tuz konsantrasyonundaki şaşırtıcı derecede büyük dalgalanmalarda kısa bir süre için hayatta kalabilirler. Birçok tür, aşağıda açıklanan fıçıyı çok tuzlu suda oluşturur; Ancak bu, aşağıda açıklanan anhidrobiyozun özel bir durumudur ve ortamın yüksek tuzluluğu ile bağlantılı iç dehidrasyona doğrudan bir yanıttır. Namlunun oluşumu bu nedenle resmi olarak bir ozmobiyotik fenomen olarak kabul edilmez.
kriyobiyoz
Kriyobiyoz, düşük sıcaklıklara dayanma yeteneğidir. Metabolizmanın neredeyse tamamen durmasıyla karakterize edilen kriptobiyozun bir tezahürüdür. Dinlenme durumu denilen bir durum oluşur . Aşağıda daha ayrıntılı olarak açıklanan varillerin oluşturulması mümkündür, ancak zorunlu değildir.
Sıcaklık yavaşça düştüğünde, enzimlerin çoğu aktivitelerini kaybettiği için metabolizma yavaş yavaş değişir ve bunun yerine şeker trehalozu ve diğer kriyokoruyucu maddeleri sentezleyen düşük sıcaklıkta aktif katalizörler etkin hale gelir : Bunlar hassas biyomembranı korur ve temel moleküllere bağlı suyun yerini alır. . Özellikle hücre dışı vücut sıvısında, daha önce tanımlanamayan, dondurucu mikroplar gibi davranan orta ağırlıktaki moleküller, küçük olduklarında donma önleyici proteinler tarafından stabilize edilen buz kristallerinin kontrollü büyümesini uyarır.
Bu sayede hayvanların donma noktasının çok altındaki sıcaklıklarda hayatta kalmaları mümkün olur. Bu yetenek, yalnızca ortam sıcaklıklarının düzenli olarak donma noktasının altına düştüğü kutup bölgelerinin ve yüksek dağ buzullarının kolonizasyonuna izin verir. Örneğin, Echiniscus sigismundi groenlandicus alt türü , kış buzunda donmuş halde 6 ila 8 ay hayatta kalır ve gelgitlerden geçişte düzenli olarak çözülme ve donmayı tolere eder .
Tardigrades, boş uzayda on gün bile hayatta kaldı. FOTON-M3 uydusu ile ESA , 2007 yılında uzayda birkaç tardigrad örneğini vakuma, soğuğa ve UV radyasyonuna maruz bıraktı. Döndüklerinde, bilim adamları en aşırı koşullara maruz kalan tardigradlar arasında bile hayatta kalanları buldular.
Tardigradlar, yeterince yavaş soğurlarsa -273 santigrat derece gibi aşırı sıcaklıklarda hayatta kalabildikleri için, dünya dışı yaşam formları olduklarından şüphelenilmektedir . Bu sıcaklıklara doğal ortamlarında asla uzaktan bile ulaşılamadı ve bu yetenek doğal seleksiyon yoluyla yeryüzünde ortaya çıkmış olamazdı . Bu görüş, yeryüzündeki donma sıcaklıklarının altında, dokusundaki suyu trehaloz gibi koruyucu maddelerle değiştirerek ve donma sürecini kontrol ederek hayatta kalan bir canlının, daha fazla evrimsel adaptasyon olmaksızın -273 santigrat derecelik sıcaklıklarda potansiyel olarak hayatta kalabileceği gerçeğiyle karşılanmaktadır. Tardigrade, çok hücreli poliekstremofillerdir .
anhidrobiyoz
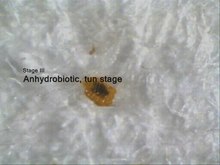
örneği kullanılarak Anhidrobiyoz .
Aşama 1: Susuz kaldığında hala hidratlı ve aktif.
Aşama 2: 10 dakika sonra bacaklar kurur ve içeri çekilir.
Aşama 3: varil aşaması (anhidrobiyoz).
Aşama 4: su ilavesinden sonra eski haline getirme.
Anhidrobiyoz, ağır su kaybı yoluyla vücudun dehidrasyonundan kurtulma yeteneğidir. Hemen hemen tüm karasal türlerde bulunur ve silindirik, hareketsiz direnç aşamaları olan varillerin oluşumu ile ilişkilidir . Anhidrobiyotik durumdaki varillerde metabolizma tespit edilemediğinden, anhidrobiyoz ayrıca kriptobiyoz genel terimi altına girer.
Bazı türlerde, çok sayıda birey gerçek namlu oluşumundan önce toplanır ve gevşek bir top oluşturur. Bu kümelenme davranışsal bir adaptasyon olarak görülür ve muhtemelen içerideki hayvanları çevresel etkilerden biraz daha fazla korur, böylece dehidrasyon süreci onlar için biraz daha yavaş olur - çok hızlı dehidrasyon ayrıca tardigratta ölüme neden olabilir.
Namlu oluşumu morfolojik reaksiyonlarla başlar: bacaklar içeri çekilir ve genel vücut yüzeyi büyük ölçüde azalır. Diğer şeylerin yanı sıra mantar saldırılarına karşı koruma sağlaması gereken yağda çözünen maddeler olan lipidler , kütiküldeki gözeneklerden giderek daha fazla salınır . Bu lipidlerdeki bir faz değişikliğinin bir sonucu olarak , kütikül belirli bir zamanda aniden su geçirmez hale gelir. Bu, suyun buharlaşmasını azaltır ve hazırlama süresinin, koruyucu bileşiklerin sentezlenmesi gereken anhidrobiyotik duruma yeniden uzatılmasına izin verir.
Anhidrobiyoz ile ilgili temel sorun, proteinler , fosfolipidler veya nükleik asitler gibi önemli makromoleküllerin yapısal bütünlüğünün her ne pahasına olursa olsun korunması gerektiğidir, aksi takdirde hücrelerde geri dönüşü olmayan hasar meydana gelir. Bu bağlantıların çoğu, gevşek şekilde bağlanmış su molekülleri ile çevrilidir ve bunların kaybı, aralarında kontrolsüz reaksiyonları tetikleyecektir. Tardigradların evrimsel tarihleri boyunca buldukları sorunun çözümü, dehidrasyon sırasında bağlı suyu, rehidrasyon sırasında (su ile ıslanma) kolayca parçalanabilen diğer bileşiklerle değiştirmektir. Bu bileşiklerin en önemlisi , anhidrobiyotik durum için hazırlık sırasında büyük miktarlarda üretilen ve bazen başlangıç durumuna kıyasla 23 kat takviye edilen şeker trehalozudur . Sadece biyomembranı korumakla ve suyu alınmış proteinler ile karbonhidratlar gibi diğer hücre bileşenleri arasındaki reaksiyonları engellemekle kalmaz, aynı zamanda önemli makromolekülleri de yok edebilecek kontrolsüz oksidasyonu önler . Yeni araştırmalar, tardigradların , susuz kaldıklarında giderek artan şekilde üretilen ve vitrifiye edilen , yani cam benzeri, koruyucu bir malzeme oluşturan yeni bir düzensiz protein tipine sahip olduğunu göstermektedir . Bu protein ve trehaloz ailesine ek olarak, gliserin de muhtemelen bu süreçlerde rol oynar . Bu arada lipit damlacıkları vücut dokularını birbirinden ayrı tutma görevine sahiptir, bu normal durumda çok yaklaşma riski taşımaz, ancak varil durumunda temas eder ve istenmeyen çapraz bağlanma reaksiyonlarını tetikleyebilir. Hayvanların normal durumlarından sabit bir varil aşamasına dönmeleri yaklaşık 5 ila 7 saat sürer.
Hayvanlar anhidrobiyotik duruma ulaşır ulaşmaz, en azından kuru bir ortamda daha fazla metabolik aktivite saptanamaz; özellikle oksijen tüketimi sıfıra düşer. Bu aşamada hayvanlar aşırı dehidrasyona, tuzlu suya, etere , saf etanol veya sıvı helyuma daldırmaya, -196 °C ile +90 °C arası sıcaklıklara, yüksek hidrostatik basınç ve vakuma dayanabilir .
Tekrar yeterli su olduğu anda, normal duruma geri dönüş, restitüsyon gerçekleşir. Bunu yapmak için trehaloz aerobik olarak, yani atmosferik oksijen kullanılarak parçalanır ve ardından su ile değiştirilir. İyileşme süresi , ortamın sıcaklığına, oksijen içeriğine ve pH değerine ve ayrıca kriptobiyotik durumda harcanan zamana bağlıdır ve ayrıca türden türe biraz farklılık gösterir. Çoğu durumda, çok uzun süreli namlu durumunda birkaç gün bile olsa, on dakika ile birkaç saat arasındadır.
Aktif bir yaşam aşamasına dönüşün başarılı olup olmadığı, öncelikle fıçı oluşumunun doğru sırasına ve hayvanların beslenme durumuna bağlıdır. Bu bağlamda özellikle önemli olan, yukarıda bahsedildiği gibi sadece kriptobiyotik durumda rol oynamakla kalmayan, aynı zamanda gliserol ve trehaloza dönüştürülebilen ve aynı zamanda restitüsyon işlemi için enerji sağlayan lipit rezervleridir. Hayvanlar bu rezervleri düzenli aralıklarla yenileme fırsatına sahip oldukları ve ayrıca herhangi bir dehidrasyon yalnızca yavaş bir şekilde gerçekleştiği sürece, düzenli aralıklarla aktif durumdan anhidrobiyotik duruma geçebilirler ve bunun tersi de geçerlidir. Örneğin, gelgit bölgesinde yaşayan Echiniscoides cinsinin bireyleri deneysel olarak altı saatlik bir döngüye sokulabilir.
Tardigrade, kriptobiyotik durumda yaşamın tanımı hakkında sorular ortaya çıkarır : metabolizma, büyüme ve üreme, genellikle canlı bir organizmanın karakteristik özellikleri olarak belirtilir. Ancak, bu özelliklerin hiçbiri fıçı aşamasında bulunmaz, bu nedenle aktif duruma dönüş, 18. yüzyılın İtalyan doğa bilimci Lazzaro Spallanzani'nin sözleriyle "ölülerden diriliş" olarak tanımlanabilir. Öte yandan, tardigradların çoğu (hepsi olmasa da) anhidrobiyotik aşamadan aktif duruma geri dönse de, tardigradolog John H. Crowe 1975'te şunu sordu:
- O halde bu, onların "ölü" iken "öldüğü" anlamına mı gelir?
- O halde bu, onların "ölü" iken "öldükleri" anlamına mı geliyor?
Crowe'un görüşüne göre, yaşamın tanımı tekil özelliklere indirgenemez; Bunun yerine, yaşayan bir sistemin yapısal sürekliliğinin ve bütünlüğünün devam eden varlığını kelimenin tam anlamıyla "hayati" olarak görür.
Üreme ve Yaşam Döngüsü
Tardigrad hem eşeysiz hem de eşeyli olarak çoğalabilir. Çoğu tür sadece cinsel olarak ürer.
Eşeysiz üreme
Eşeysiz üreme ancak partenogenez , yani daha sonra dişilere dönüşen döllenmemiş yumurtaların olgunlaşması ile mümkündür . Echiniscus ve Pseudoechiniscus cinsleri de dahil olmak üzere birçok türde bulunur . Bazı durumlarda daha sonra cüce erkekler tanımlanabilse de, erkek hayvanlar genellikle bilinmemektedir. Bu ve diğer türlerde partenogenetik ve eşeyli üremenin bir arada var olduğu belirlenebilir; erkekler temelde uygun, cinsel açıdan aktif, uygun olmayan, partenogenetik olarak üreyen dişilerden ayırt edebilirler. Partenogenez ile ilgili önemli bir avantaj, bir eş bulma çabasının olmaması, ciddi bir dezavantaj ise genetik çeşitliliğin azalmasıdır. Bununla birlikte, bunun olumsuz sonuçları, kriptobiyoz yoluyla olumsuz çevresel etkilerden basitçe kaçınma yeteneği ile kısmen dengelenir.
Eşeyli üreme
Tardigratta eşeyli üreme her zaman olmasa da çoğunlukla iki ayrı cinsiyetin varlığı ile sonuçlanır. Kendi kendine döllenme yeteneğine sahip bazı hermafrodit (hermafrodit) türler bilinmektedir; Yumurta ve sperm hücreleri aynı gonadda olgunlaşır. Onlar da bir eş aramak için herhangi bir enerji harcamak zorunda kalmama avantajına sahiptirler, ancak nihayetinde kendilerini genetik rekombinasyon olasılığı ile klonlayan partenojenik bireylerden farklıdırlar .
Bununla birlikte, türlerin büyük çoğunluğu için, dışarıdan ayırt etmek her zaman kolay olmayan ayrı cinsiyetler vardır. Nüfus yapısına ilişkin ayrıntılı çalışmaların olduğu yerlerde, hemen hemen her zaman açık bir kadın fazlalığı vardır.
Döllenme dişinin vücudunun içinde olduğu gibi dışında da gerçekleşebilir, dişiler için yumurtlama çoğunlukla bir tüy dökümü ile bağlantılıdır. Her iki sürecin ayrıntıları, diğer şeylerin yanı sıra, hayvanların yaşam alanlarına bağlıdır.
Deniz türlerinde, erkeğin spermi çoğunlukla dişilerin seminal veziküllerinde birikir, daha sonra yumurtalarını serbestçe bırakır ve alg hücreleri gibi substrata bağlanır. Ek olarak, dişinin, döllenmemiş yumurtalarını, reddedilen kütikülde tüy döktükten hemen sonra bırakması ve daha sonra erkek tarafından harici olarak döllenmesi de olur.
Limnoterrestrial türlerde, sperm bazen dişinin tüy dökümü sırasında veya öncesinde eski ve yeni deri arasındaki boşluğa enjekte edilir; dişi, atılan tüy dökme gömleğine yumurtalarını bırakır bırakmaz, döllenme gerçekleşir. Özellikle karasal türlerde, erkeğin bir dişiye tutunduğu çiftleşme sıklıkla görülür; Böyle bir bağlantı, cinsel eylemin dış rahatsızlıklarında bile terk edilmez. Erkek, dişinin tüy dökümünü bitirmeden önce spermini dişinin genital yoluna sokar. Alternatif olarak, sperm ayrıca seminal veziküllerde de bırakılabilir veya doğrudan dış deriden vücut boşluğuna enjekte edilebilir; ikinci durumda döllenme dişi gonadda gerçekleşir.
Bırakılan yumurta sayısı türe göre 1 ile 35 arasında değişmekte olup, bu sayı yaş ilerledikçe sürekli artmaktadır. Bir ömür boyunca, bekar dişiler 100'den fazla yumurta üretebilir. Deniz türlerinde çevresel koşullara bağlı olarak ince veya kalın kabuklu iken, karada yaşayan türlerde farklı yumurta şekilleri gözlemlenebilir: Düz yumurtalar çoğunlukla dişilerin tüy döken gömleklerinde, kalın, özenle süslü ve muhtemelen dehidrasyona dayanıklı kabuklar yosun bitkilerinde serbesttir veya kabuk parçaları yapıştırılmıştır. Bazı tatlı su türleri, depo olarak böceklerin veya diğer eklembacaklıların döken dış iskeletlerini kullanır.
Üreme sırasında özel davranışsal uyarlamalar sadece nadir durumlarda bulunur: bazı türlerde ilkel bir “çiftleşme ritüeli” gözlemlenmiştir. Erkek dişiyi kafasındaki cirri ile okşar. Bir süre sonra, bu şekilde uyarılan dişi, yumurtalarını bir kum tanesinin üzerine bırakır ve erkek, bunun üzerine spermini bırakır. Bazı dişilerin yumurtladıkları dökülen gömlekleri bir süre yanlarında sürüklemeleri bazen basit bir "kuluçka bakımı" vakası olarak yorumlanır.
Koşullar uygunsa, genç yaklaşık 5 ila 40 gün sonra yumurtadan çıkar. Bunu yapmak için genişleyen vücutları yumurta kabuğunu patlatana kadar ortamdan sıvı emerler. Bazı türler ayrıca yumurta kabuğunu kırmak için ağız stiletlerini kullanır. Tüm süreç genellikle sadece birkaç dakika sürer.
Genç hayvanlar çoğunlukla boyasızdır ve yetişkinlerden daha az kıl, cirri veya pençeye sahiptir. Aksi takdirde, bunlara çok benzerler, böylece gelişimleri doğrudan gerçekleşir, yani. larva aşaması olmadan. Heterotardigrada sınıfından bazı türlerde anüs ve genital açıklık ancak bir veya iki tüy dökümünden sonra oluşur; bu bazen dolaylı bir gelişmenin göstergesi olarak yorumlanır. Çoğu durumda genç hayvanlar, hücre sayısını artırarak değil, yalnızca bireysel hücre hacmini artırarak büyür, böylece birçok doku yumurtadan çıktıktan sonra zaten yetişkin formunun son hücre sayısını gösterir. Bununla birlikte, mitoz (hücre bölünmeleri) ara sıra , muhtemelen ölü hücreleri değiştirme amacına hizmet eden sonraki aşamalarda hala tespit edilebilir. Her durumda, cinsel olgunluğa ancak birkaç tüy dökümünden sonra ulaşılır.
Tardigradın normal ömrü üç ay ile iki buçuk yıl arasındadır; çoğu deniz türünün gerçek ömrüne karşılık gelir. Bununla birlikte, limnotrestrial türlerde, hayvanların yaşamı, etkilenen bireylerin yaşlanmadığı kriptobiyotik durumlar tarafından bazen veya sıklıkla kesintiye uğrar. Bu şekilde, yosun yaşayan türler genellikle dört yıl veya daha fazla gerçek bir ömre ulaşır; bireysel durumlarda da onlarca yıl sürebilirler. Bir botanik müzesinin kurumuş yosununda, 120 yıl sonra susuz bir halde "canlandırılabilecek" bir tardigrad keşfedildi.
Tardigrad ve adam
Tardigrad, neredeyse tüm insan habitatlarında bulunabilir, ancak küçük boyutları ve olağandışı yaşam tarzları nedeniyle pek fark edilmezler. Acil bir ekonomik, tıbbi veya veterinerlik önemi olmadığı için çoğu insan tarafından bilinmezler. Onları ilk kez görenler genellikle onları “sevimli” olarak tanımlar, saygın zoolojik yayınlarda bulunabilen ve muhtemelen yalnızca ayı şeklindeki vücutlarından değil, aynı zamanda hayvanların beceriksiz hareketlerinden de ilham alan bir sıfattır. . Bunları monoksenik bir kültürde, yani en az bir başka türle birlikte tutmak mümkündür.
Tehlike hakkında bilgi mevcut değildir; ancak bir tür olan Thermozodium esakii'nin soyu tükenmiş olabilir. Tardigrade, çevresel toksinlere , özellikle kükürt trioksite karşı çok hassas olduğundan , hayvanları bir yerin çevresel kalitesinin göstergesi olarak kullanmak için fikirler vardır. Macar araştırmacılar tarafından yosunlardan kaynaklanan ağır metal kirliliğinin Tardigrada'ya verdiği zarar üzerine çalışmalar sunuldu. Kentsel habitatlarla ilgili çalışmalar henüz mevcut değildir.
kabile tarihi
Modern formlar
| İlgili taksonların temsilcileri |
|---|


.jpg)
Tardigratın en yakın akrabaları iki çok farklı grupta bulunabilir: Kabuklular (Crustacea), araknidler (Arachnida), kırkayaklar (Myriapoda) ve böcekleri (Insecta) içeren eklembacaklılar , en yaygın hayvan filumlarıdır, kolomb Bacaklı solucanlar olarak tanımlanabilecek (Onychophora), oldukça belirsiz bir takson oluşturur. Tardigradlar, eklembacaklılar ve kütükler, Panarthropoda olarak bilinen monofiletik takson olarak adlandırılan doğal bir aile grubunu birlikte oluştururlar. Bu grubun tüm hayvanlarında meydana gelen vücut segmentasyonu ve eşleştirilmiş vücut uzantılarının görünümü, ortak türetilmiş bir özellik olarak görülebilir; kütikülün yapısı ayrıca üç takson tarafından da paylaşılır.
Panarthropoda içinde, tardigradlar geleneksel olarak güdük ayaklarla birlikte protoarthropodlar olarak gruplandırılır:
| panartropoda |
|
||||||||||||
Başlangıçta, fikir, kolomb ve tardigradın henüz eklembacaklıların tam organizasyon düzeyine ulaşmadığı bir rol oynadı. Bununla birlikte, evrim teorisi açısından, az ya da çok gelişmiş formlar arasındaki ayrımın artık modası geçmiş olduğu düşünülmektedir; modern sınıflandırmalar yalnızca taksonların birbirleriyle olan gerçek filogenetik ilişkilerini yansıtmayı amaçlar.
Protoarthropoda'nın ortak özellikleri, hem columbus hem de tardigrade'de bulunabilen kese benzeri vücut uzantıları, güdük bacaklarıdır. Ayrıca bir dizi başka homoloji de vardır , yani ortak bir öncü yapıya kadar izlenebilen özellikler. Özellikle, muhtemelen üç bölümden oluşan tardigrad kafa, büyük olasılıkla kolobus gövdesinin ilk üç birimine karşılık gelir. Stilettoları buna göre ikinci segmentin güçlü bir şekilde değiştirilmiş vücut uzantıları olarak görülebilir ve bu nedenle kadife solucanların çenelerine homolog olacaktır. Pençe yapısı da tardigrade ve columbus'ta çok benzer. Bu çarpıcı benzerliklere rağmen, Protoarthropoda'nın doğal bir aile grubu oluşturup oluşturmadığı tartışmalıdır, çünkü muhtemelen belirtilen özelliklerin tümü semplesiomorflardır , yani tüm Panarthropoda'nın atalarında zaten bulunurlar. 21. yüzyılın başında, bunun yerine tardigrad ve eklembacaklılar arasında bir kardeş-grup ilişkisi tercihi ortaya çıktı:
| panartropoda |
|
||||||||||||
Tardigratın diğer ilişkisi geleneksel olarak annelid solucanlarda (Annelida) görülür . Onlarla, diğer şeylerin yanı sıra, son sertleştirilmiş "pençeler" ve vücudu öne doğru kapatan bir terminal ağız pozisyonu ile yumuşak, eklemsiz vücut uzantılarını paylaşırlar. Fransız doğa bilimci Georges Cuvier'e kadar uzanan bu Articulata hipotezi, 21. yüzyılın başında hala birçok destekçisine sahipti, ancak 1990'ların sonlarına doğru morfolojik ve moleküler genetik bulgular nedeniyle giderek daha fazla sorgulandı. Bunun yerini bir grup solucan benzeri hayvanda, Cycloneuralia'nın Panarthropoda'nın en yakın akrabalarını sağladığı bir kavram aldı: Bu taksonda iplik (Nematoda) ve sicim solucanları (Nematomorpha) ve aynı zamanda oldukça belirsiz üç hayvan grubu priapulida (Priapulida ), Hookweed (Kinorhyncha) ve korse hayvanları (Loricifera). Panarthropoda gibi, tüm bu taksonların özelliği, hücresel olmayan dış derilerini veya kütiküllerini, en azından yaşam döngülerinin bireysel aşamalarında reddetmeleri; bu nedenle tüy döken hayvanlar (Ecdysozoa) olarak adlandırılırlar :
| Tüy döken hayvanlar (Ecdysozoa) |
|
||||||
soyu tükenmiş formlar
Tardigrad fosilleri son derece nadirdir ve bu nedenle bu hayvan grubunun gelişimini anlamaya çok az katkıda bulunur. Erken jeolojik çağa ait bazı buluntulara ek olarak , geç jeolojik çağdan sadece birkaç bireyin kehribar içinde korunduğu bilinmektedir .
Lobopods , bir grup solucan gibi jeolojik dönemleri hayvanların Kambriyen'den ve Ordovisiyen'den önceki kökenli Ediacarian , artan oranda görülen Tactopoda olası temsilcileri olarak, takson eklem bacaklılar ve tardigrade oluşan . Güdük ayağına benzer, eklemsiz, torba benzeri kütük bacaklar üzerinde koştular ve bu nedenle geleneksel olarak bu hayvan kabilesine atanırlar. Bazı modern kladistik çalışmalar bu değerlendirmenin temelsiz olduğunu düşünüyor ve bunun yerine lobopodları hem kadife solucanların hem de taktopodların ortaya çıktığı doğal olmayan ilişkisel bir grup olarak görüyor. Bu alternatif görüşü ifade eden önerilen bir varyant aşağıdaki şemada gösterilmektedir:
| panartropoda |
|
|||||||||||||||||||||||||||||||||||||||
Cins Aysheaia hala pek çok orijinali vardır, böylece tüm diğer Panarthropoda evrimsel kardeş grubunu stumpworts içine sonra böler oluşturan, uzmanlaşmamış uzun bacaklar veya göze çarpmayan vücut bölümleme eşleşmeyen bir ince yüzeysel zil basit satırda gibi özellikler ve diğer tüm gruplar ayrıldı. Ağır zırhlı Cardiodictyon , Hallucigenia veya Paucipodia gibi bir dizi lobopod , muhtemelen , bir yanda atanmamış Luolishania cinsine ve diğer yanda Kerygmachela kierkegaardi ve Tactopoda türlerinden isimsiz bir taksona bakan soyu tükenmiş bir yan dal oluşturur .
Bu hipoteze göre, Tactopoda'nın en yakın kardeş grubu olan Kerygmachela kierkegaardi, Kuzey Grönland'ın erken Kambriyen'inden bilinmektedir ve tardigrad ve eklembacaklılar gibi, dışarıdan da açıkça bölümlere ayrılmıştır. Gizemli Anomalocaris fosillerinin aynı zamanda Tactopoda'nın kök hattına mı ait olduğu veya yukarıda gösterildiği gibi, eklembacaklılarla tardigrattan daha yakından ilişkili olup olmadığı açık değildir .
Tardigrada açıkça atanabilen ilk fosiller Sibirya Kuonamka Formasyonu'ndan gelmektedir. 530 milyon yıllık Orta Kambriyen kalkerinde , vücut şekli ve boyutlarına göre tardigrad olarak tanımlanabilecek dört birey korunmuştur. Henüz tanımlanmamış türlerin, görünüşe göre, bir çift eşit olmayan pençe ile biten ve günümüz hayvanlarınınkine çok benzeyen bir kütikül yapısına sahip olan üç, hatta dört çift bacağa sahipti. Zaten çok özelleşmiş olan hayvanlar , muhtemelen modern sınıflardan birine, heterotardigrada'ya atanabilir .
Sağlam lobopoddan milimetre altı boyutlu tardigrada geçişin nasıl gerçekleştiği bilinmiyor. Olası bir mekanizma , gonadların embriyonik gelişim sırasında erken oluştuğu ve bu nedenle cinsel olgunluğun ilk durumdan daha erken gerçekleştiği bir süreç olan progenez olacaktır . Bu, vücudun daha fazla gelişmesini ve farklılaşmasını durdurur , böylece yetişkin hayvanda önemli ölçüde küçük bir boyut gibi larva veya genç özellikler ortaya çıkar, bu fenomen pedomorfoz olarak adlandırılır .
Tardigrade ilk kez karasal habitatları fethettiğinde bile, fosil kayıtlarının olmaması nedeniyle kesin olarak söylenemez. Tardigrad, günümüzde yosun yastıklarında sıklıkla bulunabildiğinden ve yosunlar büyük olasılıkla araziyi kolonileştiren ilk bitkiler olduğundan ve dehidrasyon riski açısından tardigradların kendileri ile benzer zorluklara maruz kaldıklarından, yosunların ve tardigradların bitki örtüsünden etkilenmiş olmaları çok olasıdır. gerekli ayarlamalar ile birlikte karaya adım.
En geç bu noktada, hayvanlar kriptobiyoz olarak bilinen kuraklık dönemlerine benzersiz adaptasyonlarını geliştirmiş olmalılar. Metabolizmalarını geçici olarak kapatarak olumsuz çevresel koşullardan kaçınmalarına izin verdi. Bu, en azından çevresel seçilimin yalnızca nispeten zayıf olması sonucunu doğurur ve böylece fiziksel görünümün, fenotipin çok yavaş, Bradytelian evrim oranını açıklar .
Bu nedenle, Kretase döneminden elde edilen geç tardigrad fosillerinin, günümüz türlerine kıyasla vücut şeklinde neredeyse hiç değişiklik göstermemesi şaşırtıcı değildir . Örneğin, Kanada kehribarından elde edilen bir tür olan Beorn leggi, Eutardigrada sınıfına zaten atanabilir; aynı bölgeden kötü korunmuş isimsiz bir genç Heterotardigrada sınıfına ait olabilir. ABD'nin New Jersey eyaletinden, muhtemelen Turoniyen denilen geç Kretase döneminden kalma başka kehribar fosilleri bilinmektedir . Onlar da muhtemelen Eutardigrada'ya atanmışlardır ve bu gelişme çizgisinin 65 milyon yıldan fazla bir süredir var olduğu sonucuna varılmasına izin vermezler.
Diğer tardigrad fosilleri, Antarktika'nın altındaki turba bataklıklarında hayatta kalan 7000 yıldan biraz daha eski yumurtalardır . Farklı tardigrad türlerinin farklı sıcaklık ve nem tercihleri olduğundan, o zamanki iklimi belirlemek için bitki polenine benzer şekilde bataklığın farklı katmanlarında görünen yumurtaların kullanılmasına ilişkin hususlar vardır.
sistematik
Tardigratların monofiletik bir takson oluşturduğuna , yani ortak bir ana türe geri döndüğüne ve bu türün tüm torunlarını içerdiğine dair ciddi bir şüphe yoktur . Önemli sinapomorfiler , ortak, türetilmiş özellikler, örneğin, teleskopik olarak geri çekilebilir bacaklar ve ağız stilesidir.
2005 yılına kadar 160 deniz formu dahil olmak üzere yaklaşık 930 tür tanımlanmıştır. Gerçek tür sayısı doğal olarak bilinmemekle birlikte 10.000 civarında olduğu tahmin edilmektedir. Ek olarak, morfolojik kriterlere göre sınırlandırılan birçok “tür”, muhtemelen taksonun biyolojik çeşitliliğini daha da artıracak olan, yalnızca moleküler genetik ile ayırt edilebilen kriptik tür gruplarını gizlemektedir.
Aralarındaki ilişkiler hala belirsiz olan üç farklı sınıf vardır:
- Gibi heterotardigrada tüm türler aslında bir geri yok olsa bile "zırhlı" su ayıları denilen ve bireysel zırh plakaları (skleritler) bölünmüş manikür içine sertleşir. Birçok formda, cirri ve clavae gibi göze çarpan uzantılar gözlemlenebilir; bacaklar hem pençelerde hem de yapışkan disklerde bitebilir. Genital açıklık her zaman doğrudan vücut yüzeyindedir, boşaltım ve osmoregülasyon için Malpigh bezleri oluşmaz. Heterotardigrada hem deniz hem de karasal habitatlarda bulunur.
- Ötardigrad kişinin "çıplak" su ayıları dediği gibi , derileri incedir ve sertleşmez. Kafada göze çarpan duyusal kıllar bu grupta hiçbir zaman bulunmaz; bacaklar temelde pençelerle biter. Heterotardigradanın aksine, fallop tüpü rektuma açılır ve böylece bir kloak haline gelir; Özel Malpigh bezleri atılım için kullanılır. Bazı deniz türleri de mevcut olmasına rağmen, Eutardigrada'nın çoğu tatlı suda veya karada yaşar.
- Mesotardigrada , resmi olarak Thermozodiidae familyasında yer alan Thermozodium esakii adlı tek bir kayıp türden bilinmektedir. Açıklamaya göre, kafasında yanal bir sirrus vardır, ancak klava yoktur; ağız dört siğil benzeri çıkıntı (papilla) ile çevrilidir. Bacağın tabanında ayrıca papilla bulunur, bacağın ucu 6 ila 10 basit pençeye dönüşür; Malpigh bezleri bulunur. Türler içinde yosun yastıkları üzerinde bulundu , sıcak bahar tipi , yerellik Japon kenti yakınlarında Nagasaki'ye , ancak İkinci Dünya Savaşı sonrasında bir depremle yıkıldı. Bugüne kadar hiçbir tip materyali korunmadığından ve türe rastlanmadığından, varlığı artık şüpheli olarak kabul edilmekte ve buna göre isim dubium olarak adlandırılmaktadır .
Genellikle tek varsayılmıştır sırası Heterotardigrada, ağırlıklı olarak deniz Arthrotardigrada, diğer Heterotardigrada sipariş Echiniscoidea biçimleri ve özellikle limnotardigrada orada hangi En ilkel tardi türü kapsamaktadır aile diğer çevirmek bu önce, Echiniscidae geliştirdik türleri esas olarak tatlı suda ve karada bulunan tardigrade sınıfı Eutardigrada:
| Tardigrad |
|
||||||||||||||||||||||||
Ancak bu hipotez henüz doğrulanmadı; ön moleküler genetik veriler, hem hetero- hem de öterdigradanın doğal olarak ilişkili olduğunu göstermektedir:
| Tardigrad |
|
||||||||||||
Araştırma geçmişi
Tardigradoloji veya tardigrad araştırması, 18. yüzyıla kadar uzanır . Alman doğa bilimci Johann Conrad Eichhorn , muhtemelen 10 Haziran 1767'de hayvanları gözlemleyen ilk kişiydi. Quedlinburg papazı Johann August Ephraim , keşfini ancak 1775'te ve dolayısıyla yıllar sonra Gdansk ve çevresindeki sularda bulunan ve çıplak gözle görülemeyen en küçük su hayvanlarının doğal tarihine katkılar adlı çalışmasında yayınladığından beri . Goeze bugün çoğunlukla grubun kaşifi olarak geçerlidir . İlk gözleminin 10 Aralık 1772'de hayvanlarla ilgili açıklamasını kendi yazdığı notlarda Fransızca'dan tercüme ettiği ve 1773 dergisinde yayınlanan Bay Karl Bonnets'in Insektologie'den incelemeleri olduğunu iddia edebilir . Orada, diğer şeylerin yanı sıra şunları yazdı:
"Bu küçük hayvan tuhaftır çünkü vücudunun tüm yapısı olağanüstü ve tuhaftır ve dış biçimiyle ilk bakışta küçük bir ayıya çok benzer. Bu aynı zamanda ona küçük su ayısının adını vermem için beni motive etti. […] Ancak, görünmez dünyanın bu yırtıcılarını düşünmekten korkmamak gerekir. "
Bir yıl sonra, ilk kez, İtalyan doğa bilimci Lazzaro Spallanzani'nin "ölümden diriliş" olarak tanımladığı anhidrobiyotik durumdan dönüş gözlemlendi . 1776'da Modena'da yayınlanan Opuscoli di Fisica animale e vegetabile adlı kitabının ikinci cildinde gruba Il Tardigrada adını veren de odur . 1790'da hayvanlar , İsveçli doğa bilimci ve sistematist Carl von Linné'nin Systema Naturae adlı çalışmasına dahil edildi .
İlk bilimsel monografi 1840'ta ortaya çıktı, dokuz yıl sonra ilk deniz tardigradı keşfedildi; Ancak ilk fosil türü olan Beorn leggi'nin bilimsel açıklaması 1964 yılına kadar bir asırdan fazla beklemek zorunda kaldı.
Hayvanların sistematik konumu baştan belirsizdi; Bütün sırasında 19. yüzyıla, onlar taksonomik isteğe ile rotiferler (Rotatorlar) ya da deniz örümcekleri (Pycnogonida) gruplandırılmış veya eklembacaklılar sağlanan (Arthropoda). 20. yüzyılın ilk yarısında bile , kesin filogenetik ilişkileri tartışmalı olmaya devam etti, böylece sonunda İtalyan tardigradolog Giuseppe Ramazzotti grubu kendi başına bir hayvan filumu seviyesine yükseltti. Taksonun daha ileri sınıflandırması, 1929'da tüm tardigradları hetero ve ötardigrada alt gruplarına ayıran Alman biyolog Ernst Marcus'a kadar uzanır; 1937'de Gilbert Rahm , Mesotardigrada'yı üçüncü grup olarak tanıttı .
Çeşitli
- İsrailli Ay'a iniş yapan Beresheet'in başarısız inişi, birkaç bin tardigradın dağılmasına neden oldu. Oksijensiz yapabildikleri ve donmuş halden ısındıktan sonra hayata döndürülebildikleri için bu ay görevi için seçildiler. “Ölü malzeme gibi, kurumuş halde yıllarca hayatta kalabilirler. Aydaki kazadan da sağ çıkmaları gerekirdi, ”dedi Nova Spivack, ABD dergisi Wired'a güvence verdi . Nova Spivack, “Arch Mission” vakfının direktörüdür.
Edebiyat
- EE Ruppert, RS Fox, RD Barnes: Omurgasız Zoolojisi. İşlevsel bir evrimsel yaklaşım. 7. baskı. Brooks / Cole, Londra 2004, ISBN 0-03-025982-7 , s. 510.
- RC Brusca, GJ Brusca: Omurgasızlar. 2. Baskı. Sinauer Associates, Sunderland Mass 2002, ISBN 0-87893-097-3 , s. 469.
- IM Kinchin: Tardigradların biyolojisi. Portland Press, Londra 1994, ISBN 1-85578-043-7 , s. 186.
- M. Blaxter, B. Elsworth, J. Daub: İhmal edilmiş bir hayvan filumunun DNA taksonomisi: beklenmedik bir tardigrad çeşitliliği. İçinde: Londra Kraliyet Cemiyeti Bildirileri. B Serisi. Londra 271.204, s. 189. ISSN 0375-0434
- GE Budd: "Opabinia regalis"in morfolojisi ve eklembacaklı kök grubunun yeniden yapılandırılması. İçinde: Lethaya. Taylor & Francis, Oslo 29.1996, ISSN 0024-1164 , s.
- RA Dewel, WC Dewel: Tardigradların eklembacaklıların evrimindeki yeri. İçinde: RA Fortey, RH Thomas (Ed.): Arthropod Relationships. (= Sistematik Birliği Spesifikasyonu Cilt 55). Chapmann ve Hall, Londra 1998, ISBN 0-412-75420-7 , s. 109.
- RA Dewel, DR Nelson, WC Dewel: "Echiniscus veridissimus" Peterfi'nin beyni, 1956 (Heterotardigrada). Filogenetik konumu ve eklembacaklı kafasının evrimini anlamanın anahtarı. İçinde: Linnean Topluluğunun Zooloji Dergisi. Blackwell, Oxford 116.1996, ISSN 1096-3642 , s.35 .
- JR Garey, DR Nelson, LY Mackey, L. Li: Tardigrade filogenisi, Morfolojik ve moleküler kanıtların uyumu . İçinde: Zoologischer Anzeiger. Elsevier, Jena 238.1999, ISSN 0044-5231 , s. 205.
- A. Hejnol, R. Schnabel: Ötardigrad Thulinia stephaniae belirsiz bir gelişime ve erken blastomer ablasyonlarını düzenleme potansiyeline sahiptir. İçinde: Geliştirme. 132.2005, s. 1349.
- A. Jörgensen, R. Kristensen: Tardigrada'nın Moleküler Filogenisi - Heterotardigrada'nın monofisinin araştırılması. İçinde: Moleküler Filogenetik ve evrim. Elsevier, Amsterdam 32.2004, 2, ISSN 1055-7903 , s. 666.
- DR Nelson: Tardigrada. İçinde: JH Thorp, AP Covich (Ed.): Kuzey Amerika Tatlı Su Omurgasızlarının Ekolojisi ve Sınıflandırılması. Academic Press, San Diego Ca 2001, ISBN 0-12-690647-5 , s. 527.
- DR Nelson: Tardigrade'nin Mevcut Durumu. Evrim ve Ekoloji. İçinde: Bütünleştirici ve Karşılaştırmalı Biyoloji. Lawrence 42.2002, ISSN 1540-7063 , s.652 .
- DR Nelson, NJ Marley: Lotic Tardigrada'nın biyolojisi ve ekolojisi. İçinde: Tatlı Su Biyolojisi. Blackwell, Oxford 44.2000, ISSN 0046-5070 , s.93 .
- Joh. August Ephraim Goeze: Bay Karl Bonnet'in insektolojiden incelemeleri. (PDF; 32,7 MB). Çevirmen JAE Goeze tarafından yapılan ek, Halle 1773. (Orijinal metin: Bu küçük hayvan tuhaftır, çünkü vücudunun tüm yapısı olağanüstü ve tuhaftır ve dış biçimi, ilk bakışta, en çok küçük bir ayıyı andırır. s. 367–375)
İnternet linkleri
- Tardigrada Günün Gökbilim Resim ait NASA (6 Mart den 2013 İngilizce )
- Tardigrada Bülteni (İng.)
- Tardigrades: fotoğraflar ve videolar (İng.)
- Tardigrade hakkında kapsamlı gerçek metinler, fotoğraf galerileri ve kısa filmler
- Cins seviyesine kadar taksonomi
- Mikroskopi İngiltere
- Tardigrade ile uzayda araştırma
- Tardigratların ekolojisi, fizyolojisi ve evrimsel biyolojisi için İsviçre araştırma merkezi
- Son zamanlarda, tardigrades'in DNA'larının altıda birini mikroplardan aldığı iddiası çözülmeye başlıyor (İng.)
- Claudia Ruby: Tardigrade - Sevimli hayatta kalma sanatçısı Bavaria 2 Radio bilgisi . 25 Temmuz 2019'da yayın (podcast)
- Ama yok edilemez değil - bu tardigrade'in zayıf noktası
- Lars Fischer: Mavi parıltı tardigradeyi radyasyondan koruyor , tarih: 15 Ekim 2020'den itibaren Spektrum.de
- Peter Dockrill: Tardigrades'in Temelde Yok Edilemez Olmak İçin Kullandığı Başka Bir Hile Bulduk , tarihinde: 14 Ekim 2020 bilim uyarısı
Bireysel kanıt
- ↑ Andreas Hejnol, Ralf Schnabel: Eutardigrade Thulinia stephaniae belirsiz bir gelişime ve erken blastomer ablasyonlarını düzenleme potansiyeline sahiptir. İçinde: Geliştirme. 132, 2005, sayfa 1349-1361. doi: 10.1242 / dev.01701
- ↑ Daiki Horikawa ve diğerleri .: Tardigrad Milagnes tardigradum'da radyasyon toleransı . Int. J. Radyat. Biol.82 , 2006, doi: 10.1080/09553000600972956 ( serbest tam metin ).
- ↑ Emma Brennand: Tardigrades: Uzayda su ayıları . Ed.: BBC. 17 Mayıs 2011 ( bbc.co.uk [30 Kasım 2019'da erişildi]).
- ↑ Peter Reuell: Harvard araştırması, asteroitlerin yaşamın yayılmasında kilit rol oynayabileceğini öne sürüyor. İçinde: Harvard Gazetesi. 8 Temmuz 2019, Erişim tarihi: 30 Kasım 2019 (Amerikan İngilizcesi).
- ↑ Lorena Rebecchi ve ark. : Anhidrobiyotik eutardigrade Paramacrobiotus richtersi'nin uzay uçuşuna direnci (FOTON-M3'teki LIFE-TARSE görevi) , içinde: Journal of Zoological Systematics and Evolutionary Research 49, 21 Nisan 2011, s. 98-103, doi: 10.1111 / j. 1439-0469.2010.0606.x .
- ↑ Neil A. Campbell ve ark. : Biyoloji. 8., güncellenmiş baskı. 2011, ISBN 978-3-8273-7287-1 .
- ↑ D. Schulze-Makuch ve ark. : Tardigradlar: Çok Hücreli Ekstremofillere Bir Örnek. İçinde: Joseph Seckbach ve diğerleri: Polyextremophiles - çoklu stres biçimleri altında yaşam. Springer, Dordrecht 2013, ISBN 978-94-007-6487-3 , s. 597-607.
- ↑ Thomas C. Boothby, Hugo Tapia, Alexandra H. Brozena, Samantha Piszkiewicz, Austin E. Smith: Tardigrades , Desiccation'dan Kurtulmak için Kendinden Düzensiz Proteinleri Kullanıyor . İçinde: Moleküler Hücre . kaset 65 , hayır. 6 , 16 Mart 2017, ISSN 1097-2765 , s. 975-984.e5 , doi : 10.1016 / j.molcel.2017.02.018 , PMID 28306513 ( cell.com [erişim tarihi 20 Haziran 2017]).
- ↑ D.D. Horikawa, T. Kunieda et al. : Aşırı toleranslı tardigrad Ramazzottius varieornatus'un yetiştirme sisteminin kurulması: astrobiyoloji için yeni bir model hayvan. İçinde: Astrobiyoloji. Cilt 8, Sayı 3, Haziran 2008, sayfa 549-556, doi : 10.1089 / ast.2007.0139 , PMID 18554084 .
- ↑ Fumihisa Ono et al. : Ultra yüksek basıncın küçük hayvanlar, tardigradlar ve Artemia üzerindeki etkisi Cogent Physics 3, 2016, doi: 10.1080 / 23311940.2016.1167575 (serbest tam metin).
- ↑ B. Vargha, E. Otvös, Z. Tuba: Biyoindikatör olarak yosunlu su ayıları (Tardigrada) tarafından Macaristan'da ağır metal kirliliğinin ekolojik etkileri üzerine araştırmalar. In: Ann Agric Environ Med. 2002; 9 (2), s. 141-146. PMID 12498580
- ↑ Klaus J. Müller, Dieter Waloszek, Arcady Zakharov: 'Orsten' tipi fosfatlanmış yumuşak bütünlük koruması ve Sibirya'daki Orta Kambriyen Kuonamka Formasyonu'ndan yeni bir kayıt. İçinde: Jeoloji ve Paleontoloji Yeni Yılı. Makaleler 197 (1), 1995, s. 101-118.
- ↑ J. A. E. Goeze: Bay Karl Bonnets insektolojiden incelemeler. J. J. Gebauer, Halle, 1773. Sayısallaştırılmış versiyona erişim 9 Eylül 2016. www.baertierchen.de adresinden alıntılanmıştır .
- ↑ Patrick Illinger: Tardigrade, Ay'ın ilk sakinleridir . İçinde: Süddeutsche Zeitung , 7 Ağustos 2019