
virüsler


Virüsler ( tekil : virüsü, dış teknik dil da virüsü, gelen Latince virüsü , doğal yapışkan nem, mukus, meyve suyu, [özellikle:] zehir ' ) olan bulaşıcı olarak görünür organik yapılar, viryonların dışında ait hücreler (hücre dışı) iletim yoluyla yayılır, ancak virüsler yalnızca uygun bir konakçı hücre içinde (hücre içi olarak) çoğalabilir. Sadece DNA veya RNA'dan ve bir hücreye girmelerine izin veren proteinlerden oluşurlar . Nükleik asitlerle, tüm virüsler çoğalmaları ve yayılmaları için bir "program" içerirler (bazı virüslerin ek yardımcı bileşenleri de vardır), ancak ne bağımsız bir replikasyonu ne de kendi metabolizmaları vardır ve bu nedenle bir konakçı hücrenin metabolizmasına bağlıdırlar. Bu nedenle virologlar büyük oranda virüsleri canlıların arasına dahil etmeme konusunda hemfikirdirler . Ama en azından onları “yaşama yakın” olarak kabul edebiliriz çünkü genellikle kendi kopyalamalarını kontrol etme ve evrimleşme yeteneklerine sahiptirler .
2011 yaklaşık 1.8 milyon farklı, virüsler için konakçı olarak hareket ettiği bilinen mevcut organizma türleriydi , ancak yalnızca yaklaşık 9110 virüs türü . Virüsler ökaryotların ( insanlar dahil bitkiler , mantarlar ve hayvanlar ) ve prokaryotların ( bakteriler ve arkeler ) hücrelerine saldırır . Prokaryotları konakçı olarak kullanan virüslere bakteriyofaj denir; özel olarak arkelere saldıran virüsler için, ancak bazen arkeofaj terimi kullanılır.
Virüsler ve viral enfeksiyonlarla ilgilenen bilim, viroloji olarak bilinir .
Keşif tarihi
19. yüzyılın ortalarında virüs terimi yalnızca “ zehir ” veya “ miasma ” ile eşanlamlı olarak kullanılıyordu . Virüsler, 19. yüzyılın sonlarından beri yalnızca ayrı bir biyolojik birim olarak biliniyor. Viral hastalıkların tanımları, ilk tedavi yöntemleri gibi çok daha eskidir. Gönderen Mezopotamya orada etrafında 1780 M.Ö. yasal metindir. Muhtemelen kuduza yakalanmış köpeği bir kişiyi ısırıp öldüren bir adamın cezalandırılmasıyla ilgilidir ( Codex Eschnunna §§ 56 ve 57). Beyanlar bilinmektedir gelen Mısır hiyeroglif muhtemelen bir sonuçlarını göstermek polio enfeksiyonu.
"Virüs" terimi ilk olarak MÖ 1. yüzyılda Cornelius Aulus Celsus tarafından kullanılmıştır . Kullanılmış. O tarif tükürük o iletilen kuduz "zehirli" olarak. 1882'de Adolf Mayer'in tütün mozaik hastalığı ile ilgili deneylerine öncülük etti ve ilk olarak farkında olmadan enfekte olmuş bitkilerin özsuyu yoluyla sağlıklı bitkilere viral bir patojen bulaşması ( bulaşma ) ve böylece bunlarda da hastalığı tetikledi.
Bu aktarım, 18. yüzyılın başlarında virüs kelimesiyle ilişkilendirildi. Times of London , virüs bulaşmasını bir doktorun ölüm ilanında açıklıyor : Parçalanmış bir cesedi dikerken, elini bıçakladı, "bu, virüs maddesinin bir kısmını bulaştırdı ya da başka bir deyişle, ona kokuşmuşluk aşıladı" (böylece bir az virüs maddesi iletildi veya başka bir deyişle çürüme aşılandı).
Mayer'den bağımsız olarak, Dimitri Iwanowski , 1892'de yaptığı bir deneyde, tütün bitkilerindeki mozaik hastalığının, bakteri geçirmez bir filtre ( Chamberland filtresi) kullanılarak süzülerek giderilemeyen ve bu nedenle partikülleri normalden önemli ölçüde daha küçük olan bir madde tarafından tetiklenebileceğini gösterdi. bakteri gerekiyordu. Ivanovsky, hastalığın nedeninin zehir olduğundan şüpheleniyordu. Martinus Willem Beijerinck bu çelişki hâlâ özellikle hastalıktan etkilenen büyüyen edildi yapraklar görülmektedir sonra: “. Bir zehir de mevcut hücreleri nüfuz isterken, virüsler besbelli geliştirmek amacıyla hücre bölünmesini gerek"
Bir hayvan virüsünün ilk kanıtı, 1898'de şap hastalığı virüsünü keşfeden Friedrich Loeffler ve Paul Frosch tarafından yapıldı (ayrıca bkz . virolojik teşhis ). Birçok virüsün boyutu 1930'larda William Joseph Elford tarafından ultrafiltrasyon yöntemleri kullanılarak belirlendi .
Daha önce eski - dolaylı - Viral hastalık nedeniyle bir kanıtı (150 milyon yıl küçük iki ayaklı dinozor deforme kemiklerden eski Dysalotosaurus lettowvorbecki türetilmiş) ait Natural History Berlin Müzesi'nde saklanan ve belirtiler arasında Paget hastalığı kemikte paramiksovirüs enfeksiyonu var .
özellikler
Virüsler, genetik bilgi taşıyan DNA veya RNA'ya sahiptir. Ancak metabolizmaları yoktur, enerjiyi dönüştüremezler ve protein üretemezler. Ancak bu yapıların oluşumu ve çoğalmaları, belirli koşullar altında , bir canlının hücresindeki metabolizmanın işleyişi ile mümkündür. Bunun için özellikle protein sentezi için gerekli olan bilgiler virüs tarafından nükleik asidi ( DNA veya RNA ) üzerinde taşınır . Esasen bir virüs, bilgileri bir konak hücrenin metabolizmasını virüslerin yeniden ortaya çıkacağı şekilde kontrol edebilen bir nükleik asittir . Replikasyonu yapısını olduğu gibi, virüs nükleik asit, konakçı hücre içinde yer alır virüs proteinleri ile ribozom olarak sitoplazması daha donatmak Virüs parçacıkları (virionlar) hücre. Virionun herhangi bir virüs zarfının lipidleri de konakçı hücreden kaynaklanır.
Virüsler iki şekilde gelir:
- Birincisi, konakçının hücrelerinde bir nükleik asit olarak . Nükleik asit, replikasyonu ve ikinci virüs formunun yeniden üretimi için bilgileri içerir . Bu formda, genetik materyallerinin DNA olarak mı yoksa RNA olarak mı depolandığına bağlı olarak DNA virüsleri ve RNA virüsleri arasında bir ayrım yapılır .
- İkincisi, konakçı hücrelerden salgılanan ve diğer konakçılara yayılmasını sağlayan bir viryon olarak .
İlgili rezervuar konakçısındaki ve muhtemelen ara konaktaki yayılma ve etki ile ilgili olarak, virüs türleri bulaşıcılık , bulaşıcılık ve patojenite veya virülans özellikleri açısından sıklıkla birbirinden çok açık bir şekilde farklılık gösterir .
Genel olarak, RNA polimerazlarının daha yüksek hata oranı nedeniyle RNA virüsleri , DNA virüslerinden çok daha değişkendir, çünkü RNA polimerazları genellikle prova okuması yoktur - eksonükleazın işlevi vardır. Bir istisna nidovirals bir kanıt okuma işlevi vardır, exoribonuclease Ekson genom boyutu çok daha az sınırlı olduğu anlamına gelir. Yüksek mutasyon oranı nedeniyle, RNA virüsleri daha fazla kusurlu, bulaşıcı olmayan viral partiküller üretir, bu da azaltılmış işlevsellik nedeniyle uygunluk maliyetleri olarak adlandırılır . Ancak, bunlar olabilir ayrıca yeni ana veya ara ana bilgisayarlara daha çabuk adapte bir seyrinde bağışıklık kaçırma ve kaçış bağışıklık tepkisini yoluyla kaçış mutasyon .
Virionların özellikleri

Hücrelerin dışındaki bir virüs parçacığına virion (çoğulu viria, virionlar) denir . Virionlar, deoksiribonükleik asitler (DNA) veya ribonükleik asitler (RNA) gibi nükleik asitler içeren ve genellikle bir protein kapsülü ( kapsid ) içeren parçacıklardır . Ancak, bir kapsül eksik örn. B. bunun yerine bir ribonükleoproteine sahip olan influenza virüsü . Bazı virionlar, lipid çift tabakası viral membran proteinleri ile serpiştirilmiş olan bir biyomembran tarafından da sarılır . Bu virüs zarfı olarak bilinir . Geçici çoğaltma fazın başlangıcına kadar kapsid ek olarak bir virüs kılıfı olan virüsler olarak anılır saran , böyle bir zarfsız virüs olarak adlandırılır zarfsız . Bazı virionların başka ek bileşenleri vardır.
Virionların çapı yaklaşık 15 nm (örneğin Circoviridae ) ila 440 nm ( Megavirus chilensis ) arasındadır. Virionlar, bakterilerden önemli ölçüde daha küçüktür, ancak ne kapsid ne de virüs zarfı olmayan viroidlerden biraz daha büyüktür .
Protein kapsidi, örneğin ikosahedral , izometrik, sarmal veya mermi şeklinde farklı şekillere sahip olabilir .
Bir virüsün serolojik olarak ayırt edilebilen varyasyonlarına serotipler denir .
Virionlar virüsleri yaymak için kullanılır. Konakçı hücrelere tamamen veya kısmen (en azından nükleik asitlerini) nüfuz ederler (onları enfekte ederler). Bundan sonra konağın metabolizması virüs nükleik asidini çoğaltmaya başlar ve diğer virion bileşenleri üretilir.

Adenovirüs , bir virion kapsidinin modeli

Bir ikosahedral virüs kapsidinin şeması

Konik bir virüs kapsidinin simetrisi
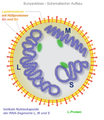
Bunyaviridae , yapının şeması

Bir lambda fajından şematik kesit (virüs ailesi Siphoviridae )

Arenaviridae yapısının şeması

Grip virüsü viryonu

HIV viriyonu
3D grafik SARS CoV-2 virüsü (renkli model)









sistematik konum
Virüsler, esasen, bir nükleik asit formunda kendi üremeleri için salt materyal programlardır. Spesifik genetik bilgilere sahip olmalarına rağmen replikasyonları için gerekli sentez ekipmanlarına sahip değillerdir . Virüslerin canlı olarak adlandırılıp adlandırılamayacağı , yaşamın tanımına bağlıdır . Genel olarak tanınan, tartışmasız bir tanım henüz mevcut değil. Çoğu bilim adamı canlı varlıklar olarak değil sınıfladıkları'nı virüsleri yok - bilimsel tartışma henüz tamamlanmış olmasa da, örneğin, genom büyüklüğü Kafeterya roenbergensis virüsü olan genomun boyutuna göre bir sınırlandırmasını bulanıklaşmaya başlıyor.
Virüsler genellikle parazitler arasında sayılmaz çünkü parazitler canlılardır. Bazı bilim adamları, virüsleri bir konak organizmayı enfekte ettikleri ve metabolizmasını kendi üremeleri için kullandıkları için hala parazit olarak görüyorlar . Bu araştırmacılar, virüsleri , en az bir genomdan oluşan ve çoğalmak için bir konak hücreye ihtiyaç duyan “zorunlu hücre içi parazitler” (her zaman hücre içinde parazit olan yaşam formları) olarak tanımlamaktadır . Canlı veya cansız olarak sınıflandırılmalarına bakılmaksızın, virüslerin davranışlarının yaygın parazitlerinkine çok benzer olduğu konusunda hemfikir olunabilir . Prionlar , işlevsiz DNA dizileri ve transpozonlar gibi virüsler bu anlamda "parazit" olarak tanımlanabilir.
Çarpma ve yayma

1. Veziküler stomatit virüsü
2. HIV
3. Vaccinia virüsü
4. Herpes simpleks virüsleri
5. Bakterilerdeki gibi fagositoz (resim) 6. Simian virüsü 40

Bir virüsün kendisi herhangi bir metabolik işlem yapamaz, bu nedenle çoğalmak için konakçı hücrelere ihtiyaç duyar . Çoğaltma çevrimi bir viryon, bir virüsten, genel olarak başlar ataşeliğini (adsorpsiyon) a yüzey proteini virüs bir konakçı hücre üzerinde bu kullanan bir şekilde reseptöre . Bakteriyofajlar söz konusu olduğunda, bu, genetik materyalinin bir hücreye enjekte edilmesiyle yapılır ; ökaryotlar söz konusu olduğunda , viryonlar endositoz tarafından tersine çevrilir ve daha sonra endozom zarına nüfuz eder , örn. B. Füzojenik bir protein tarafından . Absorpsiyondan sonra, bir virion replikasyondan önce ilk olarak zarflarından (kaplamadan) arındırılmalıdır. Virüsün genetik materyali, nükleik asidi daha sonra konak hücrede kopyalanır ve zarf proteinleri ve muhtemelen virionların diğer bileşenleri de virüs genomunun genleri kullanılarak konak hücre tarafından sentezlenir ( protein biyosentezi / gen ekspresyonu) . Böylece hücrede yeni virüsler oluşur ( morfogenez ), viryonlar halinde salınan, ya hücre zarı tarafından çözülür ( hücre lizisi , litik virüs replikasyonu) ya da parçalanarak (virüs tomurcuklanması, tomurcuklanması) tarafından boşaltılır ( salgılanır ) virüs zarfının bir parçası olarak hücre zarı . İmmünoevazinlerin yardımıyla konağın bağışıklık savunması baskılanır. Enfekte olmuş bir konak hücrede yeni oluşan virionların sayısı, patlama boyutu olarak adlandırılır .
Başka bir olasılık, virüs genomunun konakçınınkiyle ( provirüs ) entegrasyonudur . Bakteriyofaj lambda gibi ılıman virüslerde durum böyledir .
Virüs replikasyonunun konak hücre üzerindeki etkisine sitopatik etki (CPE), tüm konak organizma üzerindeki etkisine viral hastalık veya viroz denir . Farklı sitopatik etki türleri vardır: hücre lizizi , piknoz ( poliovirüsler ), hücre füzyonu ( kızamık virüsü , herpes simpleks virüsleri , parainfluenza virüsü ), intranükleer kapanımlar ( adenovirüsler , kızamık virüsü), intraplazmik kapanımlar ( kuduz virüsü , çiçek virüsleri ).
Virüsler birçok şekilde yayılabilir. Örneğin, insanlara patojen olan virüsler, damlacık enfeksiyonu (örn. grip virüsleri) yoluyla hava yoluyla veya yayma enfeksiyonu (örn. herpes simpleks ) yoluyla kontamine yüzeyler yoluyla bulaşabilir . Bitki virüsleri genellikle böcekler veya iki bitki arasındaki mekanik bulaşma yoluyla veya tarımda kontamine aletler yoluyla bulaşır. Teorik biyolojide virüslerin ve diğer patojenlerin epidemiyolojik kinetiğinin soyut bir görünümü geliştirilmiştir .
evrim
Menşei
Virüslerin kaynağı bilinmiyor. Günümüzde çoğu araştırmacı virüslerin hücresel yaşamın öncüleri olmadığını , canlılardan kopan canlıların genleri olduğunu varsaymaktadır . Temelde iki farklı yaklaşımın olduğu çeşitli olasılıklar hala tartışılmaktadır:
- Virüsler çok ilkeldir; ilk hücreden önce ve en ilkel yaşam biçimlerini bile üreten o kimyasal “ilkel çorba” da ortaya çıktılar ; RNA genomları ile DNA öncesi dünyadan kalmadırlar. Bu yaklaşım, örneğin, Félix Hubert d'Hérelle (1924) ve Salvador Edward Luria (1960) tarafından temsil edildi.
- Virüsler, oluştuklarında zaten var olan eksiksiz organizmalarda bir tür düşüştür.
Bundan, üç teori formüle edilmiştir.
- Kendi kendini kopyalayan moleküllerden türetilmiştir (birlikte evrim). Bu teori, virüslerin kökeninin ve evriminin, kendi kendini kopyalayabilen en basit moleküllerden başladığını varsayar. O zaman bu tür bazı moleküller sonunda hücre olarak görülebilecek organizasyonel birimler oluşturmak için bir araya gelecekti. Aynı zamanda diğer moleküller de kendilerini hücrelere paralel olarak gelişmeye devam eden ve onların paraziti haline gelen virüs parçacıklarına paketlemeyi başardılar.
- Dejenerasyon yoluyla virüs gelişimi ( parazit ). Bu teori, ilk virüslerin başlangıçta bakteriler (veya varsayımsal ribositler ) gibi serbest yaşayan organizmalardan ortaya çıktıklarına ve sonunda genetik bilgilerinin yavaş yavaş ve sürekli olarak kaybolduğuna dair yukarıda sunulan ikinci olası yaklaşıma dayanmaktadır. kendilerine kaybolan işlevleri sağlamak için bir konak hücreye güvenen hücre parazitleri. Bu bağlamda dikkati artan bir kavram (ki Virozelle English virocell ): Bir virüsün asıl fenotipi enfekte hücre, virion (virüs partikülleri), ancak üreme veya dağılımın sadece bir aşaması, polen veya sporlara benzer .
- Konakçı hücrenin kendi RNA veya DNA moleküllerinden virüs oluşumu. Araştırma için en olası görünen bu üçüncü teori, virüslerin doğrudan konak hücrenin RNA veya DNA moleküllerinden ortaya çıktığını belirtir. Virüslerin genetik materyali olarak, bağımsız hale gelen bu nükleik asitler, konak hücrenin genomundan veya RNA'sından bağımsız olarak çoğalma yeteneği kazanmış, ancak sonunda parazit olarak kalmışlardır ( S. Luria , 1960). Olası geçiş formlarının örnekleri, transpozonlar ve retrotranspozonlardır .
değişkenlik
Bir virüsün (veya herhangi bir genin) evrimi için değişkenliği ve seçimi önemlidir. Değişkenlik (tüm organizmalarda olduğu gibi) genetik materyalin replikasyonundaki kopyalama hatalarından kaynaklanır ve diğer şeylerin yanı sıra bağışıklık kaçırma ve konak spektrumunu değiştirmek için kullanılırken , seçim genellikle konağın (bağışıklık) tarafından gerçekleştirilir. cevap.
Daha yüksek organize canlılar geliştirdik çok etkili bir olasılık genetik değişkenliği ile rekombinasyon ve çaprazlamadan içinde , cinsel üreme özellikle çevreye uyum yönünde ve ilgili türler, böylece daha da geliştirilmesi. Virionlar veya virüsler, hayatta kalabilen ve üremeleri için canlı konakçılara bağımlı olan ve bu nedenle de eşeyli üreme olmaksızın sadece mutasyona uğrama yetenekleriyle yayılan yapılar olarak , genetik değişkenlik için en azından eşit fırsat gösterirler .
O zaman, virüslerin genomundaki bu mutasyonların, esas olarak, konakçı hücreler içinde replikasyon sırasındaki kopyalama hatalarına dayanması, sonuçta önemsizdir. Önemli olan tek şey, türlerin korunmasından kaynaklanan uyum yeteneğindeki aşırı artışın olumlu etkisidir. Bu tür hatalar, örneğin oldukça gelişmiş bir memeli hücresinde hücre ölümüne yol açabilirken, virüsler için aslında büyük bir seçim avantajına sahiptirler .
Replikasyondaki kopyalama hataları nokta mutasyonlarda , yani rastgele gen konumlarında yanlış bazların dahil edilmesinde ifade edilir . Virüsler, daha gelişmiş hücrelerin aksine, çok az onarım mekanizmasına sahip olduklarından veya hiç tamir etmedikleri için bu hatalar düzeltilmez.
Virüslerdeki özel genetik değişim biçimleri, örneğin influenza virüsleri durumunda, antijen kayması ve antijen kayması (genetik yeniden sınıflandırma ) terimleri kullanılarak burada ayrıntılı olarak açıklanmaktadır.
Konak reaksiyonları
Virüs enfeksiyonu, konakçılarında çeşitli savunma tepkileri oluşturur . Kullandıkları için virüsler, sadece hücre içinde çoğaltılır gerekli yapı blokları ve enzimler arasından sitozolü bir konakçı hücre için çoğaltma . Bu nedenle, kısıtlama veya direnç faktörleri olarak bilinen çeşitli hücre içi savunma mekanizmaları ortaya çıkmıştır . Birlikte bakteriler de dahil olmak üzere CRISPR ve sınır enzimleri bakteriyofajların önlemek için bir hücre içinde kullanımı bulunmaktadır ökaryotlarda bu tür. B. myxovirus direnç faktörü Mx1 , PAMP reseptörleri , dsRNA aktif inhibitörü ve çeviri DAI , melanom farklılaşma antijeni 5 ( MDA-5 ), bir oligoadenilatm OAS1 sentaz , Langerin , tetherinin , SAM alan ve HD 1. Alanın protein ( SAMHD1 ), RIG-I , APOBEC3 , TRIM5alfa , protein kinaz R ve RNA interferansı .
Hayvanlarda, özellikle omurgalılarda, ek bir bağışıklık tepkisi gelişmiştir. Kısmen doğuştan , kısmen kazanılmış . Edinilmiş veya edinilmiş bağışıklık tepkisi sırasında, virüsün tek tek bileşenlerini ( antijenler ) bağlayabilen antikorlar ve sitotoksik T hücreleri oluşturulur . Bu, virüsleri ve virüs bulaşmış hücreleri tanımalarını ve yok etmelerini sağlar.
birlikte evrim
Doğal konaklardaki patogenezde bir gözlem, konakçıya adapte olmuş patojenlerin , kendi gelişimleri için ihtiyaç duydukları ve bağışıklık sistemi hücre hasarı ve apoptoz ile aktive edildiğinden , konakçıya genellikle çok fazla zarar vermediğidir . Bir bağışıklık reaksiyonundan kaçınmak , diğer konakçılara replikasyon ve iletimi ( iletim ile eş anlamlıdır ) kolaylaştırır . Bazı virüsler ömür boyu vücutta kalır. Reaktivasyon, semptomlar olmasa bile zaman zaman ortaya çıkabilir. (Ayrıca bakınız: Patojen kalıcılığı .) Örneğin, herpes simpleks virüsleri , daha az belirgin semptomlarla Alman nüfusunun %90'ından fazlasının enfeksiyon oranlarına ( kontaminasyon ile eş anlamlıdır ) ulaşır . Maymun bağışıklık eksikliği virüsü üretmez AIDS doğal konaklarda aksine, HIV insanlarda. Buna karşılık, insanlarda Ebola virüsü ile olan enfeksiyonlar , doğal konaklarında değil, etkili bulaşma gerçekleşmeden önce yüksek virülanları nedeniyle bazen kendi kendine söner , çünkü konak ciddi şekilde zayıflar ve kısa sürede ölür, dolayısıyla hareket alanı ve dolayısıyla hareket alanı ve dolayısıyla virüsün yayılması sınırlı. Ölüm oranı yüksek olan ciddi bir enfeksiyon seyri (bkz. öldürücülük ve ölüm oranı ), genellikle neden olan patojenin, rezervuar konakçı olarak söz konusu organizmaya henüz adapte olmadığının bir göstergesidir. Bununla birlikte, yüksek düzeyde replikasyona (ve neden olunan hasara) sahip patojenlerden kalıcı bir enfeksiyon oranına (enfekte et ve devam et , hasardan kaçınma) geçiş akıcıdır. Başka bir deyişle, uyarlanmış bulaşıcı nesneler kalıcı olma ve düzenlenmiş bir üreme hızı eğilimi gösterirken, daha az uyarlanmış patojenler enfeksiyon zincirinin erken sonlanmasına yol açma eğilimindedir . İstisnalar örn. B. Kuşlarda H5N1 virüsleri , Yersinia pestis ve insan çiçek hastalığı virüsleri . Bununla birlikte, patojenler kendi türdeşleri ile rekabet halinde olduğundan ve daha az üreme patojeni daha çabuk yok olacağından, adaptasyon genellikle konak tarafından gerçekleşir. Bu nedenle patojenlerde patojenitede bir azalma, öncelikle artan üreme oranı ile bağlantılı olarak meydana gelir.
Konağın patojene adaptasyonu, konak kısıtlaması veya direnci olarak adlandırılır. Bilinen antiviral ve antibakteriyel mekanizmalar arasında, ökaryotlarda, insanlarda, örneğin, miksovirüs direnç faktörü Mx1 , PAMP reseptörleri , dsRNA ile aktive olan translasyon inhibitörü DAI, MDA5 , oligoadenilat sentaz OAS1 gibi konakçı reaksiyonları altında açıklandığı gibi , yer alır. langerin , tetherinin , APOBEC3 , TRIM5alpha ve protein kinaz R., ekleme bağışıklık tepkisi meydana gelir .
sınıflandırma
Geleneksel virüs sınıflandırması
1962'de André Lwoff , Robert W. Horne ve Paul Tournier , Carl von Linné tarafından kurulan canlı varlıkların ikili sınıflandırmasına dayanan ve aşağıdaki seviyeleri (taksonların sonları için model) içeren bir virüs taksonomisi ("LHT sistemi") tanıttı. parantez içinde):
- Virosphere ( Filum : Vira)
- Alt filum (... virüs)
-
Sınıf (biyoloji) (... ica)
-
Sipariş (... viral)
-
Aile (... viridae)
- Alt familya (... virinae)
-
Aile (... viridae)
-
Sipariş (... viral)
-
Sınıf (biyoloji) (... ica)
- Alt filum (... virüs)
Buna, ana bilgisayarlara dayalı gruplar halinde bir atama eşlik eder.
- Bakteriler ve arkeler (bakteriyofajlar / arkeofajlar tarafından istila)
- Algler , mantarlar ve protozoa
- Bitkiler ( viroidler tarafından da istila )
- Üç alt gruba sahip hayvanlar:
- omurgasızlar (omurgasızlar)
- Omurgalılar (omurgalılar)
- Her iki grubun temsilcileri
Çoğu virüs yukarıdaki dört gruptan sadece birine aittir, ancak Rhabdoviridae ve Bunyaviridae familyalarındaki virüs türleri hem bitkileri hem de hayvanları enfekte edebilir. Bazı virüsler sadece omurgalılarda çoğalır, fakat aynı zamanda omurgasızlar (bkz. vektör ), özellikle böcekler tarafından mekanik olarak bulaşır . Bir konak hücrenin eklem enfeksiyonu sırasında diğer virüslerden (mumya virüsler) gelen genlerin kullanımına dayanan virüslere virofajlar denir.
ICTV'ye göre virüs taksonomisi
Uluslararası Virüs Taksonomisi Komitesi (ICTV) aileleri içine eşit bir şekilde bölünmesi sağlamak için bir sınıflandırma sistemi geliştirmiştir. Dokuzuncu ICTV raporu, virüs türlerini hiyerarşik bir dallanan virüs takson sistemindeki en düşük takson olarak tanımlamaktadır .
2017 yılına kadar, taksonomik yapı temel olarak seviye düzeni ve altındaki (yukarıya bakın) geleneksel virüs sınıflandırmasıyla aynıydı ve 2018'de aşağıdaki gibi (LHC sisteminden farklı ad sonları ile) daha ileri seviyelerle desteklendi:
- Alan (tr. Bölge) ( ... viria )
- Alt-alan (tr. Subrealm) ( ... vira ) (LHC sisteminde Subphylum ile olduğu gibi, ikinci en üst seviye olarak biter)
-
Reich (tr. Kingdom) ( ... virae )
- Unterreich (tr. Subkingdom) ( ... virites )
-
Gövde veya filum ( ... viricota ) ( ... arkeotaya benzer şekilde - LHC sisteminden farklı olarak, birkaç virüs filumu mümkündür)
- Alt filum ( ... virikotina )
-
Sınıf ( ... viricetes )
- Alt sınıf ( ... viricetidae )
- Sipariş ( ... viral )
- Subordinasyon ( ... virineae )
- Aile ( ... viridae )
- Alt familya ( ... virinae )
- Cins veya cins ( ... virüs )
- Alt cins veya alt cins ( ... virüs )
- Türler veya türler ( ... virüs )
- Alt cins veya alt cins ( ... virüs )
- Cins veya cins ( ... virüs )
- Alt familya ( ... virinae )
- Aile ( ... viridae )
- Subordinasyon ( ... virineae )
- Sipariş ( ... viral )
- Alt sınıf ( ... viricetidae )
-
Sınıf ( ... viricetes )
- Alt filum ( ... virikotina )
-
Gövde veya filum ( ... viricota ) ( ... arkeotaya benzer şekilde - LHC sisteminden farklı olarak, birkaç virüs filumu mümkündür)
- Unterreich (tr. Subkingdom) ( ... virites )
-
Reich (tr. Kingdom) ( ... virae )
- Alt-alan (tr. Subrealm) ( ... vira ) (LHC sisteminde Subphylum ile olduğu gibi, ikinci en üst seviye olarak biter)
Şimdiye kadar izin verilen bu seviyelerden (ICTV durumu Şubat 2019) yalnızca alan, şube, alt şube, sınıf, takım, tabi olma, aile, alt aile, cins, alt cins ve türler fiili kullanımdadır. Bu kılavuzda alt türlerin , türlerin ("bakteriyel tür" gibi çeşitler anlamında) veya izolatların tanımı yoktur . Bu nedenle tüm sıraların ad sonları bileşen olarak "vir" içerir (ancak "viroid" biçiminde değildir); kısaltmalar "V" ile bitiyor, muhtemelen arkasından bir sayı geliyor (Romen değil, Arapça). İçin viroidler ve uydu olarak subviral parçacıklar , benzer bir sınıflandırma, karakterize edici bir bileşen ile kendi adı uçlarının her biri kullanılabilir.
Mart 2019 itibariyle aşağıdaki düzenlemeler eklenmiştir:
Phylum Negarnaviricota'ya :
- muviral
- Serpentoviraller
- Jingchuvirales
- mononegaviraller
- Goujianvirales
- Bunyaviraller
- artikülaviraller
Riboviria'nın diğer emirleri ve astları :
Üst sıralarda gruplanmayan siparişler:
Baltimore Sınıflandırması

Nobel ödüllü biyolog David Baltimore tarafından önerilen sınıflandırma, virüs genomunun tam biçimine ve ondan haberci RNA'nın ( mRNA ) nasıl üretildiğine dayanmaktadır. Virüs genomu, DNA veya RNA, tek sarmallı (ss) veya çift sarmallı (ds) şeklinde olabilir. Tek bir iplik orijinal olarak (İngilizce: duyu, +) veya tamamlayıcı bir biçimde (İngilizce: antisens, -) var olabilir. Belirli koşullar altında, bir RNA genomu replikasyon için geçici olarak DNA'ya dönüştürülür ( retrovirüsler ) veya tersine, bir DNA genomu geçici olarak RNA'ya kopyalanır ( pararetrovirüsler ); her iki durumda da RNA, bir ters transkriptaz (RT) ile DNA'ya geri yazılır .
Tüm virosfer aşağıdaki yedi grup tarafından tanımlanır:
- I: dsDNA virüsleri (ayrıca adenovirüsler , herpes virüsleri , dev virüsler , çiçek hastalığı virüsleri )
- II: ssDNA virüsleri (+ sarmal) DNA (artı parvovirüsler )
- III: dsRNA virüsleri ( reovirüsler dahil )
- IV: (+) ssRNA virüsleri (+ zincirli) RNA (ayrıca pikornavirüsler , togavirüsleri )
- V: (-) ssRNA virüsleri (şerit) RNA (ayrıca ortomiksovirüsler , rhabdovirüsler )
- VI: ssRNA-RT virüsleri (+ sarmal) - DNA ara aşamasına sahip RNA ( retrovirüsler )
- VII: dsDNA-RT virüsleri - RNA ara evreli DNA ( pararetrovirüsler , artı hepadnavirüsler )
Modern virüs sınıflandırmaları, ICTV ve Baltimore'un bir kombinasyonunu kullanır.
Virüs türü adlarının yazılışı
Bir virüsün resmi uluslararası, bilimsel adı, Lagos yarasa virüsü (LBV) gibi uluslararası kısaltmada da kullanılan İngilizce adıdır . Bu kısaltma Almanca'da da değişmeden kullanılır. Mantıksal olarak, Alman virüs adının Lagos-Fledermaus-Virus kısaltması da LBV'dir .
West Nile virüsü gibi İngilizce virüs adlarında normalde tire kullanılmaz ve virüs küçük harftir. Kısa çizgi İngilizce olarak yalnızca sıfatlarla , yani Kene kaynaklı ensefalit virüsü veya Avian ensefalomiyelit benzeri virüs ile görünür .
Almanca'da virüs adı kısmen tire ile yazılabilir , yani Batı Nil virüsü , hepatit C virüsü , insan herpes virüsü , Lagos yarasa virüsü , Avrupa yarasa Lyssa virüsü , bazen birlikte. Alt türlerin sayılarının önüne bir boşluk (İngilizce'de olduğu gibi) gelir ve kısaltmaların önüne bir kısa çizgi gelir, örn. B. Avrupa Yarasa Lyssa Virüsü 1 (EBLV-1), Herpes Simplex Virüsü 1 (HSV-1) ve İnsan Herpes Virüsü 1 (HHV-1).
Canlı dünyanın aksine, ICTV kurallarına göre, daha yüksek taksonların isimleri ( aile , tarikat , sınıf , filum (bölme) vb. gibi) de italik olarak yazılır. Genellikle tercüme edilmezler. , alt tiplerin isimleri ( kabileler - İngilizce suşlar anlamında ) - ve izolatlar) asla italik yazılmaz (konaklarının içerdiği herhangi bir jenerik ve / veya tür adı bile yoktur).
Sık kullanılan adlar çevrilmişse ( Coronaviridae için koronavirüsler gibi ), bu terimler italik değildir. Taksonlar genellikle yalnızca sona göre artan düzende farklılık gösterdiğinden ( Herpesviridae ve Herpesvirales gibi ), Almancalaştırılmış isimler (burada: Herpesviruses) kullanıldığında belirsizlikler sıklıkla ortaya çıkar. Bu aynı zamanda bu tür isimlerin kullanımı için de geçerlidir (örneğin, SARS-CoV-2 gibi en belirgin temsilciler için koronavirüsler ).
İnsanlar için patojenik virüsler ve neden olduğu hastalıklar

Virüsler insanlarda çok çeşitli hastalıklara neden olabilir. Ancak bu insan patojenik virüsleri burada genom ve Behüllung açısından sınıflandırılmış ve ICTV tarafından sınıflandırılmış taksonomileri listelenmiştir.
Zarflı virüsler
Çift sarmallı DNA virüsleri = dsDNA
- Aile Poxviridae
- Alt familya Chordopoxvirinae
- Cins Orthopoxvirus
- Orthopoxvirus variola = Variolavirus - çiçek hastalığı , gerçek çiçek hastalığı
- Orthopoxvirus variola var. Alastrim = kaffirpox virüsü - çiçek hastalığı , beyaz çiçek hastalığı
- Monkeypoxvirus (MPV) = Orthopoxvirus simiae - maymun çiçeği ; insanlara da aktarılabilir, semptomlar insan çiçeğinde olduğu gibi, ancak çok daha hafif
- Cins Parapoxvirus
- Parapoxvirus ovis = Orf virüsü - Orf
- Cins Molluscipoxvirus
- Molluscum Contagiosum Virüsü - Dellwarze ( Molluscum contagiosum )
- Cins Orthopoxvirus
- Alt familya Chordopoxvirinae
- Aile Herpesvirüsü
- Alt familya Alphaherpesvirinae
- Cins simpleks virüsü
- Herpes simpleks virüsü 1 (HSV-1) = İnsan herpes virüsü 1 (HHV-1) - herpes simpleks , herpes labialis , aftöz stomatit
- Herpes Simpleks Virüsü 2 (HSV-2) = İnsan Herpes Virüsü 2 (HHV-2) - Herpes simpleks , genital herpes
- Herpes B virüsü = ( Herpes virüsü simiae )
- Cins Varicellovirus
- Varicella zoster virüsü (VZV) = insan herpes virüsü 3 (HHV-3) - suçiçeği = varicella (herpes zoster), zona
- suid herpesvirus tip 1 (SHV-1) = pseudowut virüsü , Aujeszky virüsü ve diğerleri. - Aujeszky hastalığı = sözde öfke, kaşıntı salgını, deli uyuz vb. (hayvanlarda, düşük patojeniteli insanlara da aktarılabilir)
- Cins simpleks virüsü
- Alt familya Betaherpesvirinae
- Cins Sitomegalovirüs
- İnsan Sitomegalovirüsü (HCMV) = İnsan Sitomegalovirüsü (HZMV) = İnsan Herpes Virüsü 5 (HHV-5) - Sitomegali
- cins Reseolovirüs
- İnsan herpes virüsü 6 (HHV-6) - üç günlük ateş
- İnsan herpes virüsü 7 (HHV-7) - üç günlük ateş
- Cins Sitomegalovirüs
- Gammaherpesvirinae alt familyası
- cins Lenfokriptovirüs
- Epstein Barr Virüsü (EBV) = İnsan Herpes Virüsü 4 (HHV-4) - Pfeiffer glandüler ateş , Burkitt lenfoma
- Cins Rhadinovirus
- İnsan Herpes Virüsü 8 (HHV-8) - Kaposi sarkomu
- cins Lenfokriptovirüs
- Alt familya Alphaherpesvirinae
- Aile Hepadnaviridae
- Cins Orthohepadnavirus
- Hepatit B Virüsü (HBV) - Hepatit B
- Cins Orthohepadnavirus
Tek (+) zincirli RNA virüsleri = ss (+) RNA
- Aile Togaviridae
- Cins Alphavirus - arbovirüslerin etken maddesi
- Barmah Forest Virus - Grip benzeri semptomlarla Barmah Forest ateşi , salgın poliartrit
- Chikungunya virüsü (CHIKV) - Chikungunya ateşi
- Doğu At Ensefalit Virüsü (EEEV) = Doğu
- Batı At Ensefalit Virüsü (WEEV) = Batı at ensefalit virüsü - sivrisineklerden insanlara bulaşma da mümkündür (nadir!) → Batı at ensefalomiyeliti ( ensefalit / ensefalomiyelit )
- Everglades Virüsü - Everglades Ateşi
- O'nyong Nyong Virüsü (ONNV) - O'nyong Nyong Ateşi
- Mayaro ateşi virüsü (MAYV) - Mayaro ateşi
- Semliki Orman Virüsü (SFV) - Semliki Orman Ateşi
- Mucambo Virüsü - Mucambo Ateşi
- Ross River Virüsü (RRV) - Ross River Ateşi
- Sindbis virüsü (SINV) - Sindbis ateşi (eklemlerin iltihabı ["salgın poliartrit "], bazen döküntüler ve nadiren ensefalit ile birlikte )
- Cins Alphavirus - arbovirüslerin etken maddesi
- Rubivirüs cinsi
- Rubivirüs = Kızamıkçık Virüsü = Kızamıkçık Virüsü (RUBV) - Kızamıkçık
- Cins Hepacivirus
- Hepatit C Virüsü (HCV) - Hepatit C
- GB-Virus-C (hastalık değeri yok)
- Cins Flavivirüs
- Batı Nil Virüsü (WNV) - Batı Nil ateşi
- Dang virüsü (DENV) - Dang humması
- Sarı humma virüsü (YFV) - sarı humma
- Louping Ill Virus (LIV) - Louping Ill Ensefalit
- St. Louis Ensefalit Virüsü (SLEV) - St. Louis Ensefaliti
- Japonya Ensefalit Virüsü (JEV) - Japon ensefaliti
- Usutu virüsü (USUV) - ateş ve/veya döküntü gibi spesifik olmayan semptomlar
- Kyasanur Orman Hastalığı Virüsü (KFDV) - Kyasanur Orman Ateşi
- Powassan virüsü (POWV) - Powassan ensefaliti
-
TBE virüsü [İngilizce: kene kaynaklı ensefalitik virüs (TBEV)] - TBE (erken yaz meningoensefaliti)
- Avrupa / Batı kene kaynaklı ensefalit virüsü (WTBEV) alt tipi
- Alt tip Sibirya kene kaynaklı ensefalit virüsü (STBEV)
- Alt tip Uzak Doğu kene kaynaklı ensefalit virüsü (Uzak Doğu TBEV); eskiden Rus İlkbahar Yaz Ensefalit Virüsü (RSSEV) - RSSE , ayrıca RFSE (Rus İlkbahar Yaz Ensefaliti )
- Zika virüsü (ZIKV) (2 ana grup; çeşitli alt tipler) - çoğunlukla sadece deri döküntüsü, ateş, eklem ağrısı, konjonktivit
- Alt familya Orthocoronavirinae
- Cins Alfakoronavirüs
- Alt cins Duvinacovirus
- İnsan koronavirüsü 229E (HCoV-229E) - soğuk algınlığı
- Setracovirus alt cinsi
- İnsan koronavirüsü NL63 (HCoV-NL63) - soğuk algınlığı
- Alt cins Duvinacovirus
- Cins Betacoronavirus
- Alt cins Embecovirus
- Türler Betacoronavirus 1
- Alt türler insan koronavirüsü OC43 (HCoV-OC43) - soğuk algınlığı; bazen şiddetli solunum yolu enfeksiyonu, pnömoni
- Tür insan koronavirüsü HKU1 (HCoV-HKU1) - soğuk algınlığı
- Türler Betacoronavirus 1
- Alt cins Merbecovirus
- Orta Doğu solunum sendromu koronavirüsü (MERS-CoV) - grip benzeri semptomlar, şiddetli solunum yolu enfeksiyonu, zatürre ve muhtemelen böbrek yetmezliği
- Alt cins Sarbecovirus
-
SARS ile ilişkili koronavirüs (SARS-CoV) - SARS ( atipik pnömoni ), alt tipi ile
- Alt tip SARS-CoV-2 (İng. 2019-novel Coronavirus, 2019-nCoV veya Wuhan deniz ürünleri pazarı pnömoni virüsü) - COVID-19 : Alt solunum yollarının
-
SARS ile ilişkili koronavirüs (SARS-CoV) - SARS ( atipik pnömoni ), alt tipi ile
- Alt cins Embecovirus
- Cins Alfakoronavirüs
- Cins Torovirüs
- çeşitli tipler - gastroenterit
- Alt familya Orthoretrovirinae
- cins Deltaretrovirüs
- İnsan T-Lenfotropik Virüs 1 (HTLV-1) - Yetişkin T-Hücreli Lösemi , Tropikal Spastik Paraparezi
- İnsan T-lenfotropik virüsü 2 (HTLV-2) - lösemi (?)
- İnsan T Lenfotropik Virüsü 3 (HTLV-3) - bilinmiyor
- İnsan T Lenfotropik Virüsü 4 (HTLV-4) - bilinmiyor
- Cins Lentivirüs
- İnsan İmmün Yetmezlik Virüsü Tip 1 (HIV-1) - AIDS
- İnsan İmmün Yetmezlik Virüsü Tip 2 (HIV-2) - AIDS
- cins Deltaretrovirüs
Tek (-) - zincirli RNA virüsleri = ss (-) RNA
- Aile Arenaviridae
- Cins Mammarenavirus
- Alt cins LCMV / Lassa kompleksi (eski dünya arenavirüsleri)
- Lassa virüsü ( Lassa mammarenavirus / LASV) - Lassa ateşi
- Lenfositik koriomenenjit virüsü ( lenfositik koriomenenjit virüsü / Lenfositik koriomeningitis mammarenavirus / LCMV) - lenfositik koriomenenjit
- Mopeia virüsü (MOPV) - Mopeia ateşi ( bazen çok şiddetli ila ölümcül seyreden hemorajik ateş .)
- Lujo virüsü ( Lujo mammarenavirus / LUJV) - Lujo ateşi ( hemorajik ateş )
- Subgenus Tacaribe Kompleksi: (Yeni Dünya Arenavirüsleri)
- Chapare virüsü ( Chapare mammarenavirus / CHAV) - hemorajik ateş
- Tacaribe virüsü ( Tacaribe mammarenavirus / TCRV) - Hemorajik ateş
- Junin virüsü ( Arjantin mammarena virüsü / JUNV) - Junin ateşi (Arjantin hemorajik ateşi / AHF)
- Machupo virüsü ( Machupo mammarenavirus / MACV ) - Machupo ateşi (Bolivya hemorajik ateşi / yüksek ölümcül BHF.)
- Guanarito virüsü ( Guanarito mammarenavirus / GTOV) - Guanarito ateşi (Venezuela kanamalı ateşi / VHF)
- Sabia virüsü ( SPH 114.202 virüs / Brezilyalı mammarenavirus / Sabiá mammarenavirus / SABV) - Sabia ateş (Brezilya hemorajik ateşi / BzHF)
- Alt cins LCMV / Lassa kompleksi (eski dünya arenavirüsleri)
- Cins Mammarenavirus
- Aile Bornaviridae
- Cins Orthobornavirus (eski adıyla Bornavirus )
-
Borna hastalığı Virüs (İngilizce Borna hastalığı virüsü = BoDV.) - neden olan ajan Borna hastalığı insanlara nadir durumlarda devredilebilir atlar, koyunlar ve diğer memelilerde - ağır ( ensefalit )
-
Memeli Borna virüsü 1 (ingiliz. Memeli 1 orto Borna virüsü)
- Borna hastalığı virüsü 1 (BoDV-1) - atlarda, koyunlarda ve diğer memelilerde Borna hastalığının etken maddesidir, nadir durumlarda insanlara da bulaşabilir - insanlarda şiddetli ensefalitin nedeni olduğu kanıtlanmıştır.
- Borna hastalığı virüsü 2 (BoDV-2)
-
Memeli Borna virüsü 2 (ingiliz. Memeli 2 orto Borna virüsü)
- Kızıl sincap boravirüs 1 ( Alacalı sincap Bornavirüs 1 = VSBV-1 ) kızıl sincaplarda ( Sciurus variegatoides ) tespit edildi , ayrıca insanlara da bulaşabilir - potansiyel olarak ölümcül ensefalit
-
Memeli Borna virüsü 1 (ingiliz. Memeli 1 orto Borna virüsü)
-
Borna hastalığı Virüs (İngilizce Borna hastalığı virüsü = BoDV.) - neden olan ajan Borna hastalığı insanlara nadir durumlarda devredilebilir atlar, koyunlar ve diğer memelilerde - ağır ( ensefalit )
- Cins Orthobornavirus (eski adıyla Bornavirus )
- Aile Bunyaviridae - arbovirüslerin etken maddeleri
- Cins Orthobunyavirus
- Bunyamwera virüsü (serogrup)
- Batai virüsü (BATV) - grip benzeri semptomlar ve döküntüler
- California Ensefalit Virüsü (Serogrup) - Ensefalit
- cins flebovirüs
- Rift Valley Fever Virüsü (3 alt tip) - Rift Valley Fever
-
Tatarcık humması virüsü (SFNV) - Tatarcık humması = tatarcık ateşi
- Alt tip Karimabad virüsü (KARV)
- Alt tip kum sineği humması
- Cins Orthobunyavirus
- Alt tip Tahran virüsü (THEV)
- Toscana virüsü (TOSV) alt tipi - Pappataci ateşi
- Serotipler : Toskana (T), Sicilya (G) ve Napoli (K)
-
Kırım-Kongo ateşi virüsü (serogrup):
- Alt tip Kırım-Kongo kanamalı ateşi virüsü (CCHFV) - Kırım-Kongo kanamalı ateşi
- Hazara virüsü (HAZV) alt tipi - Kırım-Kongo ateşi
- Alt tip Khasan virüsü (KHAV) - Kırım-Kongo ateşi
- Hantaan virüsü (4 alt tip) - hemorajik ateş , nefrit
- Seul virüsü (serogrup) - hemorajik ateş
- Prospect Hill virüsü (2 alt tip) - hemorajik ateş
- Puumala virüsü (serogrup) - hemorajik ateş , pnömoni , nefrit
- Dobrava Belgrad Virüsü - Hemorajik Ateş
- Tula virüsü - hemorajik ateş
- Sin Nombre virüsü (serogrup) - şiddetli pulmoner ödemli hemorajik ateş
- Cins Marburg virüsü
- Victoria Gölü Marburg virüsü (serogrup) - Marburg ateşi ( hemorajik ateş )
- Cins Ebola virüsü
- Cins Influenzavirus A - Grip (grip)
- Grip virüsü A varyantı H1N1 - Grip (grip)
- Grip virüsü A varyantı H3N2 - Grip (grip)
- (Kuş) influenza virüsü A varyantı H5N1 , yüksek derecede patojenik kuş gribi virüsü ( HPAIV ) - hayvanlarda "kuş gribi", insanlara da bulaşabilir, ancak insandan insana pek bulaşmaz .
- Cins Influenzavirus B - Grip (grip)
- Grip Virüsü B / Victoria Hattı - Grip (Grip)
- Grip Virüsü B / Yamagata Hattı - Grip (Grip)
- Cins Influenzavirus C - Grip (grip)
- Cins Influenzavirus D - Grip (grip)
- Cins Avulavirüs
- İnsan parainfluenza virüsü (tip 1, 3) - soğuk algınlığı , parainfluenza
- Cins Morbillivirus
- Cins Henipavirus
- Hendra Virüsü , (eski adıyla Equines Morbillivirus ) - Pnömoni ; ensefalit
- Nipah Virüsü - Zatürre ; ensefalit
- Cins Rubula Virüsleri
- İnsan parainfluenza virüsü (tip 2, 4) - soğuk algınlığı , parainfluenza
- Kabakulak virüsü - kabakulak
- Cins Orthopneumovirus (önceden: Pneumovirus )
- İnsan Solunum Sinsityal Virüsü (HRSV) (Tip A, B) - solunum yolu enfeksiyonu , soğuk algınlığı
- cins metapnömovirüs
- İnsan metapnömovirüsü (HMPV) (tip A1 ila 2, B1 ila 2) - solunum yolu enfeksiyonu , soğuk algınlığı
- Cins Vesiculovirus
- Veziküler stomatit Indiana virüsü (VSV) - hayvanlarda stomatit vesicularis (ağız mukozasının kabarma ile iltihaplanması), insanlara da bulaşır
- cins Lyssavirus
- Kuduz virüsü (RABV) (önceden genotip 1) = kuduz virüsü - kuduz , hayvanlarda, insanlara da bulaşır
- Mokola virüsü (MOKV) (eski genotip 3) - hayvanlarda kuduz insanlara da bulaşabilir
- Duvenhage virüsü (DUVV) (önceden genotip 4) - hayvanlarda kuduz insanlara da bulaşabilir
- Avrupa bat-lyssa virüsü 1 + 2 (EBLV-1, -2) (önceki genotip 5 ve 6) - hayvanlarda kuduz insanlara da bulaşabilir
- Avustralya Yarasa Lyssa Virüsü (ABLV) (önceden genotip 7) - hayvanlarda kuduz , insanlara da bulaşır
zarfsız virüsler
Çift sarmallı DNA virüsleri = dsDNA
- Aile Adenoviridae
- cins mastadenovirüs
- İnsan adenovirüsleri AF (51 alt tip) - burun akıntısı , soğuk algınlığı , ishal
- cins mastadenovirüs
- Aile Polyomaviridae
- cins polyomavirüs
- BK polyomavirus (BKPyV) = BK virus (BKV) = polyoma virus hominis tip 1 - transplantasyondan sonra immünosupresif tedavi ile sonuçlanır, muhtemelen greft kaybı.
- JC poliomavirüsü (JCPyV) = JC virüsü (JCV) = poliyomavirüs hominis tip 2 - hücre içinde (bağışıklığı bastırıcı madde AİDS için) progresif multifokal lökoensefalopati (PML)
- cins polyomavirüs
- Aile Papillomaviridae
- cins papilloma virüsü
- Alt cins insan papilloma virüsü
- çeşitli insan papilloma virüsleri (HPV) - siğiller ;
- Kondiloma Virüsü 6 (HPV-6) - genital siğiller ; Çok seyrek: Tekrarlayan respiratuar papillomatozis ( larenkste soluk borusuna (trakea) ve akciğerlere giden çoğunlukla iyi huylu skuamöz epitelyal papillomların çoklu, tekrarlayan oluşumu .
- Kondiloma Virüsü 11 (HPV-11) - genital siğiller ; Çok seyrek: Tekrarlayan respiratuar papillomatoz (gırtlakta soluk borusuna (trakea) ve akciğerlere giden çoğunlukla iyi huylu skuamöz hücreli papillomların çoklu, tekrarlayan oluşumu).
- İnsan papilloma virüsü 16/18/30… (HPV-16 / -18 / -30…) - rahim ağzı kanseri (rahim ağzı kanseri)
- Alt cins insan papilloma virüsü
- cins papilloma virüsü
Tek sarmallı DNA virüsleri = ssDNA
- Aile Parvoviridae
-
- Alt familya Parvovirinae
-
- Cins Dependoparvovirus (diğer adıyla Dependovirus )
-
- Türler adeno-ilişkili virüs A (AAV-A)
- Adeno-ilişkili virüs 1 ila 4 (AAV-1 ila AAV-4)
- Tür adeno-ilişkili virüs B (AAV-B)
- Adeno-ilişkili virüs 5 (AAV-5)
- Cins Erythroparvovirus (aka Erythrovirus )
-
- Türler Primat eritroparvovirüs 1
Çift sarmallı RNA virüsleri = dsRNA
- Aile Reoviridae
- cins Rotavirüs
- çeşitli tipte - gastroenterit ile ishal
- Cins Coltivirus
- cins Rotavirüs
Tek (+) zincirli RNA virüsleri = ss (+) RNA
- Aile Caliciviridae
- cins norovirüs
-
Norovirüs (NV) = Norwalk Benzeri Virüs (NLV)
- GGI, GGII ve GGIV gruplarının insan norovirüsleri - kusma ishal = gastroenterit
-
Norovirüs (NV) = Norwalk Benzeri Virüs (NLV)
- cins sapovirüs
- Sapovirüs (SV) - gastroenterit
- cins norovirüs
- Aile Hepeviridae
- cins Hepevirüs
- Hepatit E Virüsü (HEV) - Hepatit E.
- cins Hepevirüs
- Aile Picornaviridae
- enterovirüs cinsi
- Poliovirüs tip 1-3 - çocuk felci
-
Coxsackievirus A / B - dan soğuk algınlığından için menenjit , pankreatit veya miyokardit , nadiren felç
- Coxsackievirus B1 (CVB-1) ila B 6 - soğuk algınlığı
- Echovirüs - ekzantema enanthemum , üst enfeksiyonları solunum yolu ( soğuk ), herpanjin , myopericarditis yenidoğanlarda, yaygın (yaygın) enfeksiyonu, kronik meningoensefalit olarak bağışıklığı zayıflamış hastalarda, menenjit , ensefalit , nadiren felç
-
enterovirüs
-
İnsan enterovirüsleri - soğuk algınlığı
- İnsan Enterovirüs 70 (EV-70) - akut hemorajik konjonktivit
- İnsan Enterovirüs 71 (EV-71) - meningoensefalit , döküntü ve poliomyelit benzeri sendrom = el-ayak-ağız hastalığı
-
İnsan enterovirüsleri - soğuk algınlığı
- cins hepatovirüs
- cins rinovirüs
-
rinovirüs
- İnsan rinovirüsleri -1 A (HRV-1 A) veya 1 B ila 100 - soğuk algınlığı
-
rinovirüs
- enterovirüs cinsi
onkovirüsler
İnsanlardaki en önemli kanserojen virüsler olan " oncovirüsler " grubu , Amerikan Kanser Derneği'nin tahminlerine göre, kanser vakalarının yaklaşık %17'sinden bile , dünya çapındaki tüm insan kanserlerinin yüzde 10 ila 15'inden sorumludur .
- Epstein Barr Virüsü (EBV)
- Hepatit B virüsü (HBV)
- Hepatit C virüsü (HCV)
- İnsan papilloma virüsü (HPV)
- İnsan T-lenfotropik virüsü 1 (HTLV-1)
- İnsan herpes virüsü 8 (HHV-8, ayrıca Kaposi sarkomu herpes virüsü, KSHV)
dev virüsler
antiviral ilaçlar
Virüsler veya viryonlar, bakterilerin aksine hücre olmadıkları için bu şekilde öldürülemezler. Bir viral enfeksiyonu ve virüs replikasyonunu önlemek veya önlemek ancak antiviraller kullanarak mümkündür . Özellikle biyokimyasal üreme süreçleri virüs tipinden virüs tipine çok farklı olabilir, bu da engelleyici veya engelleyici bir ajan bulmayı zorlaştırır.
Virüs normal hücrelerin içinde çoğaldığı ve merkezi biyokimyasal hücre mekanizmalarıyla çok yakından bağlantılı olduğu için söz konusu antiviral ajanlar kullanılmalıdır.
- viryonların konak hücrelere girmesini önlemek,
- virüs replikasyonu zararına hücre metabolizmasına müdahale veya
- hücrelerde olası bir virüs replikasyonundan sonra yeni virüslerin hücrelerden kaçışını durdurun.
Öte yandan, aranan bu aktif maddeler aynı zamanda vücut metabolizması, hücre yapısı ve/veya bir bütün olarak iç hücre metabolizması ile uyumlu olmalıdır, çünkü aksi takdirde sadece örneğin hücrelerde virüs replikasyonu çıkmaza girer. durur, ancak en kötü durumda tedavi edilen tüm organizmanın (hücre) yaşamı da devam eder.
Bu koşulların uzlaştırılması çok zor olduğundan, şimdiye kadar geliştirilen antiviral ilaçlar sıklıkla ciddi yan etki riski taşır . Bu dengeleme eylemi, tıbbı şimdiye kadar çoğunlukla çözülmemiş zor görevlerle karşı karşıya getiriyor.
Etkili antiviral ilaçların geliştirilmesi , virüslerin, bir kez bulunursa, son derece hızlı üreme döngüleri ve bu replikasyonun biyokimyasal doğası nedeniyle iyi bir şekilde yetenekli oldukları yararlı bir aktif bileşene karşı mücadele edilecek dirençlerinin geliştirilmesiyle de yoğunlaştırılır .
virüs tedavisi
Şu anda hastalıkları iyileştirmek için virüsleri kullanan tedaviler hakkında daha fazla araştırma var. Bu araştırma, kullanımını içerir viral vektörler olarak onkolitik virüs mücadele için tümörler olarak, faj tedavisi hedefli enfeksiyon ve için liziz arasında bakteri , bunlardan bazıları dirençli için antibiyotikler olarak, aşılar için profilaksisi ve tedavisi enfeksiyon hastalıklarının neden olduğu oluşturulması için, pluripotent kök hücreler veya için gen terapisi ve genetik bozukluklar .
Ayrıca bakınız
Edebiyat
- eski edebiyat
- Feodor Lynen : Virüs sorunu. İçinde: Angewandte Chemie . Cilt 51. No. 13, 1938, ISSN 0044-8249 , sayfa 181-185.
- güncel literatür
- Hans W. Doerr, Wolfram H. Gerlich (ed.): Tıbbi viroloji - virolojik hastalıkların temelleri, teşhisi ve tedavisi. Thieme, Stuttgart / New York 2002, ISBN 3-13-113961-7 .
- Walter Doerfler: Virüsler. Fischer Taschenbuch Verlag, Frankfurt a. M. 2002, ISBN 3-596-15369-7 .
- Dietrich Falke , Jürgen Bohl ve diğerleri : hasta başı virolojisi: klinik, teşhis, tedavi. Springer, Heidelberg ve diğerleri 1998, ISBN 3-540-64261-7 . (referanslarla birlikte)
- Matthias Eckoldt Virüsü: Parçacıklar, Paranoya, Salgınlar . Ecowin, Salzburg / Münih 2021, ISBN 978-3-7110-0275-4 .
- Dietrich Falke, Jürgen Podlech: Virüsler. İçinde: Peter Reuter: Springer Lexicon Medicine. Springer, Berlin ve ark. 2004, ISBN 3-540-20412-1 , s. 2273-2282.
- SJ Flint, LW Enquist, VR Racaniello (Ed.): Virolojinin İlkeleri. 2. baskı, ASM Press, Washington DC 2003, ISBN 1-55581-259-7 .
- Alfred Grafe: Virüsler - yaşam alanımızın parazitleri. Springer, Berlin / Heidelberg / New York 1977, ISBN 3-540-08482-7 .
- David M. Knipe, Peter M. Howley, et al. (Ed.): Fields' Virology. 2 cilt, 5. baskı, Lippincott Williams & Wilkins, Philadelphia 2007, ISBN 978-0-7817-6060-7 (virolojide standart çalışma).
- Arnold J. Levine : Virüsler: Hırsızlar, Katiller ve Korsanlar. Spektrum Akademik Yayınevi, Heidelberg 1992, ISBN 3-86025-073-6 .
- Susanne Modrow, Dietrich Falke , Uwe Truyen: Moleküler Viroloji. Biyologlar ve tıp uzmanları için bir giriş (= spektrum ders kitabı ). 2. baskı, Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1086-X .
- Stephen S. Morse: Virüslerin Evrimsel Biyolojisi. Raven Press, New York 1994, ISBN 0-7817-0119-8 .
- Sven P. Thoms: Yaşamın kökeni: Dünya'da yaşam nasıl ve ne zaman ortaya çıktı? ... (= Fischer cep kitapları; Fischer kompakt. ). Fischer Taschenbuch Verlag, Frankfurt a. M. 2005, ISBN 3-596-16128-2 .
- Luis P. Villarreal: Virüsler ve Yaşamın Evrimi. ASM Press, Washington 2005, ISBN 1-55581-309-7 .
- Ernst-Ludwig Winnacker : Virüsler: Gizli yöneticiler. Grip, AIDS ve hepatit dünyamızı nasıl tehdit ediyor? Eichborn, Frankfurt a. M. 1999, ISBN 3-8218-1598-1 .
- Gottfried Schuster: Çevredeki Virüsler. Teubner, Stuttgart 1998, ISBN 3-519-00209-4 .
- Dorothy H. Crawford: Görünmez düşman: virüslerin doğal tarihi. Oxford University Press, Oxford 2002, ISBN 0-19-856481-3 .
- Brian W. Mahy: Viroloji sözlüğü. Elsevier, Amsterdam 2008, ISBN 0-12-373732-X .
- Susanne Modrow: Virüsler: Temel Bilgiler, Hastalıklar, Tedaviler. [Tıbbi meslekten olmayanlar için kolay anlaşılır bir giriş]. Beck, Münih 2001. ISBN 3-406-44777-5 .
- Hartwig Klinker: Virüs enfeksiyonları. In: Marianne Abele-Horn (Ed.): Antimikrobiyal Tedavi. Bulaşıcı hastalıkların tedavisi ve profilaksisi için karar desteği. (Werner Heinz, Hartwig Klinker, Johann Schurz ve August Stich işbirliğiyle) 2., gözden geçirilmiş ve genişletilmiş baskı, Peter Wiehl, Marburg 2009, ISBN 978-3-927219-14-4 , s. 297-307.
- Marilyn J. Roossinck: Virüsler! Yardımcılar, düşmanlar, afiyet olsun - 101 portrede . Springer, Berlin 2018, ISBN 978-3-662-57543-7 .
- Sunit K. Singh (Ed.): Viral Enfeksiyonlar ve Küresel Değişim. [Küreselleşme ve iklim değişikliğinin virüslerin, özellikle tropik virüslerin yayılması ve bulaşması üzerindeki etkisi üzerine]. Wiley-Blackwell, Hoboken NJ 2014, ISBN 978-1-118-29787-2 (baskı); ISBN 978-1-118-29809-1 (e-Kitap).
- Eugene V. Koonin, Tatiana G. Senkevich, Valerian V. Dolja: Antik Virüs Dünyası ve hücrelerin evrimi. İçinde: Biyoloji Doğrudan. 19 Eylül 2006, Cilt 1, Madde 29, doi: 10.1186 / 1745-6150-1-29 , PMID 16984643 , PMC 1594570 (serbest tam metin) (İngilizce).
İnternet linkleri
- Virüsler: yapı, spesifik özellikler, gelişme, hücre biyolojisi, bakterilerden farklılaşma
- Uluslararası Virüs Taksonomisi Komitesi
- Virüsler nasıl çalışır?
- Virusworld (X-ray yapı analizlerinden elde edilen virüslerin 3 boyutlu gösterimleri)
- Virüsler insan evrimini nasıl teşvik etti?
Bireysel kanıt
- ^ Karl Ernst Georges : Kapsamlı Latin-Almanca özlü sözlük . 8., geliştirilmiş ve artırılmış baskı. Hahnsche Buchhandlung, Hanover 1918 ( zeno.org [21 Ocak 2020'de erişildi]).
- ↑ Çevrimiçi Düden: Virüs, şu ya da bu
- ↑ Karin Mölling : Yaşamın Süper Gücü . Virüslerin muhteşem dünyasına seyahat edin. 1. baskı, Beck, Münih 2015, ISBN 978-3-406-66969-9 .
- ↑ Atık sularda on binlerce bilinmeyen virüs . Tarih: ssinexx.de, 6 Ekim 2011, en son 17 Eylül 2014'te erişildi.
- ↑ Uluslararası Virüs Taksonomisi Komitesi (ICTV): Virüs Taksonomisi: 2020 Yayını. - EC 52, çevrimiçi toplantı: Ekim 2020; E-posta onayı: Mart 2021. Şu adreste : talk.ictvonline.org ; en son 17 Mayıs 2021'de erişildi.
- ↑ TA McAllister ve diğerleri.: Ruminant Beslenme Sempozyumu : Ruminant metanogenezisi azaltmak için stratejileri belirlemek için genomik ve transkriptomik kullanımı. İçinde: ACCESS DL. 2015 Arşivlenen kopyası ( hatırasısın içinde 7 Nisan 2016 Internet Archive ) doi: 10,2527 / jas.2014-8329
- ↑ Shmoop Biology: Phages Shmoop University, 2016.
- ↑ Pierer'in Geçmişin ve Bugünün Evrensel Sözlüğü . 4. baskı. Verlagbuchhandlung von HA Pierer , Altenburg 1865 ( zeno.org [21 Ocak 2020'de erişildi] ansiklopedi girişi “Virüs”).
- ^ Rahip Dr. Peckwell In: The Times, 23 Ağustos 1787, s. 2.
- ^ Matthias Eckoldt: VİRÜS - Parçacıklar, Paranoya, Salgınlar . Ecowin, Salzburg 2021, ISBN 978-3-7110-0275-4 , s. 61 .
- ↑ Florian Witzmann ve ark.: Jurassic bir dinozorda kemiğin Paget hastalığı. İçinde: Güncel Biyoloji. Cilt 21, Sayı 17, R647-R648, 2011, doi: 10.1016 / j.cub.2011.08.006 ( tam metin PDF dosyası olarak ).
- ^ R. Sanjuan, MR Nebot, N. Chirico, LM Mansky, R. Belshaw: Viral Mutation Rates . İçinde: Viroloji Dergisi . 84, No. 19, 2010, ISSN 0022-538X , s. 9733-9748. doi : 10.1128 / JVI.00694-10 .
- ↑ JW Drake, JJ Holland: RNA virüsleri arasındaki mutasyon oranları. İçinde: Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildiriler Kitabı. 1999, Cilt 96, Sayı 24, sayfa 13910-3. PMID 10570172 , PMC 24164 (serbest tam metin).
- ↑ Donald W. Klein, Lansing M. Prescott, John Harley: Mikrobiyoloji . Wm.C. Brown, Dubuque, Iowa 1993, ISBN 0-697-01372-3 .
- ↑ MA Martinez ve diğerleri: RNA Virüslerinin Quasispecies Dynamics . İçinde: G. Witzany (Ed.): Virüsler: Yaşamın Temel Ajanları . Springer, 2012, ISBN 978-94-007-4898-9 , sayfa 21-42.
- ↑ C Lauber, JJ Goeman, C Parquet Mdel, P Thi Nga, EJ Snijder, K Morita, AE Gorbalenya: RNA virüslerindeki en büyük genom genişlemesinde genom mimarisinin ayak izi . İçinde: PLoS Pathog . Cilt 9, Sayı 7, Temmuz 2013, sayfa E1003500. doi : 10.1371/journal.ppat.1003500 .
- ↑ DA Steinhauer, JJ Holland: RNA virüslerinin hızlı evrimi. İçinde: Mikrobiyolojinin Yıllık İncelemesi. 1987, Cilt 41, sayfa 409-33. PMID 3318675 .
- ↑ John B Carter, Venetia A Saunders: Virology: Principles and Applications. 1. baskı, Wiley, Chichester UK 2007, ISBN 0-470-02387-2 , s. 6.
- ^ Matthias G. Fischer, Michael J. Allen, William H. Wilson ve Curtis A. Suttle: Dikkat çekici bir gen tamamlayıcısı olan dev virüs, deniz zooplanktonunu enfekte eder . İçinde: Ulusal Bilimler Akademisi Bildiriler Kitabı . 2010. doi : 10.1073 / pnas.1007615107 .
- ^ Salvador Edward Luria, James E. Darnell : Genel Viroloji. 3. baskı, John Wiley & Sons, New York ve ark. 1978, ISBN 978-0-471-55640-4 .
- ↑ Luis P. Villarreal, Günther Witzany: Virüsler, hayat ağacının köklerinde ve gövdesinde bulunan temel ajanlardır. İçinde: Teorik Biyoloji Dergisi. Cilt 262, No. 4, 2010, sayfa 698-710, doi: 10.1016 / j.jtbi.2009.0.014 .
- ↑ P. Forterre: virocell kavramı ve çevresel mikrobiyoloji . İçinde: ISME Dergisi . Cilt 7, 2012, s. 233-236. doi : 10.1038 / ismej.2012.110 . PMC 3554396 (ücretsiz tam metin).
- ↑ David M. Needham, Susumu Yoshizawa, Toshiaki Hosaka ve diğerleri: Dev virüslerin belirgin bir soyu, tek hücreli deniz yırtıcılarına bir rodopsin fotosistemi getiriyor . İçinde: PNAS. 23 Eylül 2019, doi: 10.1073 / pnas.1907517116 , ISSN 0027-8424 ( PDF ).
- ↑ Georg Löffler, Petro E Petrides (Ed.): Biochemie und Pathobiochemie (= Springer ders kitabı. ) 7., tamamen gözden geçirilmiş baskı, Springer, Berlin / Heidelberg / New York 2003, ISBN 3-540-42295-1 .
- ↑ David Moreira, Purificación López-García: Virüsleri hayat ağacından dışlamak için on neden. İçinde: Doğa İncelemeleri Mikrobiyoloji . Cilt 7, Nisan 2009, s. 306-311, doi: 10.1038 / nrmicro2108 .
- ^ A b V. J. Torres, DL Stauff ve diğerleri: Konak heme yanıt veren ve virülansı modüle eden bir Staphylococcus aureus düzenleyici sistem. İçinde: Hücre konakçısı ve mikrop. 19 Nisan 2007, Cilt 1, Sayı 2, sayfa 109-19, PMID 18005689 , PMC 2083280 (serbest tam metin).
- ↑ G. Silvestri: Doğal olarak SIV ile enfekte olmuş isli mgenauylar: neden AIDS geliştirmediklerini anlamaya daha yakın mıyız? İçinde: Tıbbi Primatoloji Dergisi. 2005, Cilt 34, Sayı 5-6, sayfa 243-52, PMID 16128919 .
- ↑ MJ Pantin-Jackwood, DE Swayne: Kuşlarda kuş gribi virüsü enfeksiyonunun patogenezi ve patobiyolojisi. In: Revue scientifique et tekniği (Uluslararası Epizootik Ofisi). 2009, Cilt 28, No. 1, sayfa 113-36, PMID 19618622 .
- ↑ KD Mir, MA Gasper, V. Sundaravaradan, DL Sodora: Doğal konaklarda SIV enfeksiyonu: akut-kronik geçiş fazı sırasında immün aktivasyonun çözülmesi. İçinde: Mikroplar ve Enfeksiyon. 2011, Cilt 13, No. 1, sayfa 14-24, PMID 20951225 , PMC 3022004 (serbest tam metin).
- ↑ MJ. Adams, EJ. Lefkowitz, AM. Kral, EB. Carstens: Yakın zamanda Uluslararası Virüs Sınıflandırma ve Adlandırma Kodunda üzerinde anlaşmaya varılan değişiklikler . İçinde: Viroloji Arşivleri . Cilt 158, Sayı 12, Aralık 2013, sayfa 2633-9. doi : 10.1007 / s00705-013-1749-9 . PMID 23836393 .
- ↑ Uluslararası Virüs Taksonomisi Yürütme Kurulu , Virüs Taksonomisi: 2018 Yayını , Virüs ve tür adları nasıl yazılır
- ↑ Uluslararası Virüs Taksonomisi Komitesi Yürütme Komitesi : Virüs sınıflandırmasının yeni kapsamı: virosferi 15 hiyerarşik kademeye bölmek. İçinde: Doğa Mikrobiyolojisi. Cilt 5, sayfa 668-674, 27 Nisan 2020, doi: 10.1038 / s41564-020-0709-x ; ve Nadja Podbregar: Virosphere için bir soy ağacı. Tarih: ssinexx.de , 29 Nisan 2020. Her iki makale de Ocak 2020, i. H. Detaylar Usta Türler Listesi Mart 2020 ICTV ait (MSL) sayılı 35 henüz dikkate alınmamıştır. Ancak ICTV'nin temel amacı için bunun bir anlamı yoktur, MSL # 35 ile gelişim sadece belirtilen yönde devam etmiştir.
- ↑ Uluslararası Virüs Taksonomisi Komitesi (ICTV): ICTV Ana Tür Listesi 2018b.v2 (MSL # 34v)
- ↑ Susanne Modrow, Dietrich Falke, Uwe Truyen: Moleküler Viroloji. 2. baskı, Spektrum - Akademischer Verlag, Heidelberg / Berlin 2003, ISBN 3-8274-1086-X .
- ↑ Th. Mertens, O. Haller, H.-D. Klenk (Hrsg.): Viral hastalıkların teşhisi ve tedavisi - viroloji için toplumun kılavuzları. 2. baskı, Elsevier / Urban & Fischer, Münih 2004, ISBN 3-437-21971-5 .
- ↑ Thomas Berg, Norbert Suttorp: Bulaşıcı Hastalıklar. Thieme, Stuttgart 2004, ISBN 3-13-131691-8 .
- ↑ ICTV: Virüs, tür ve diğer takson adları nasıl yazılır
- ↑ Tıbbi Laboratuvar Teşhisi Sözlüğü . S. 402-403: Barmah Orman Virüsü (BFV).
- ^ Gerhard Dobler, Horst Aspöck: İnsan enfeksiyonlarının etken maddeleri olarak sivrisinekler tarafından bulaşan arbovirüsler . İçinde: Horst Aspöck (ed.): Eklembacaklılardan hasta (= Denisia. Cilt 30). Biyoloji Merkezi (Linz'deki Yukarı Avusturya Devlet Müzeleri), 2010, ISSN 1608-8700 , burada s. 518: Barmah Forest-Fieber. - 2.6.4. Klinik ( PDF olarak tam metin ).
- ↑ Hastalık Kontrol ve Önleme Merkezleri (CDC): Doğu At Ensefaliti . Tarih: cdc.gov, 5 Nisan 2016; en son 30 Ağustos 2016 tarihinde erişildi.
- ↑ Adriana Delfraro, Analía Burgueño, Noelia Morel u a. .: Batı At Ensefalitinin Ölümcül İnsan Vakası, Uruguay. İçinde: Gelişen Bulaşıcı Hastalıklar. Cilt 17, Sayı 5, Mayıs 2011, s. 952–954 → Harfler, doi: 10.3201 / eid1705.101068 ( PDF dosyası olarak tam metin ).
- ↑ Araştırma Kapısı, Mopeia virüsünün Afrika arena virüsüne karşı varlığı. Kasım 2011, erişim tarihi 23 Mayıs 2019 .
- ↑ Robert Koch Enstitüsü - Broşür: Borna Hastalığı Virüsü 1 ile enfeksiyonlardan kaçınma hakkında bilgi 1. Açık: rki.de ; 5 Aralık 2020 tarihinde erişildi.
- ↑ Federal Eğitim ve Araştırma Bakanlığı: Enfeksiyon araştırması - tarla fareleri ölümcül Borna virüsünü bulaştırabilir. Tarih: bmbf.de 8 Ocak 2020, en son 5 Aralık 2020'de erişildi.
- ↑ Bernd Hoffmann, Dennis Tappe, Dirk Höpe ve diğerleri: Ölümcül İnsan Ensefaliti ile İlişkili Alacalı Bir Sincap Bornavirüsü. İçinde: New England Tıp Dergisi. 2015, Cilt 373, sayfa 154-16, doi: 10.1056 / NEJMoa1415627 .
- ↑ MSD El Kitabı: Tekrarlayan solunumsal papillomatoz (laringeal papillomlar). Açık: msdmanuals.com Şubat 2019 itibariyle; en son 29 Nisan 2021'de erişildi.
- ↑ NCBI: Dependovirus (cins)
- ↑ D. Martin, JS Gutkind: İnsan tümörü ile ilişkili virüsler ve kanserin moleküler mekanizmalarına dair yeni görüşler . İçinde: Onkogen . Cilt 27, Sayı 2, 2008, sayfa 31-42. PMID 19956178 .
- ↑ C. Zimmer: Kanser - evrimin bir yan etkisi mi? İçinde: Bilim Spektrumu. 9, 2007, s. 80-88.
- ^ M. Stadtfeld ve diğerleri: Viral Entegrasyon Olmadan Üretilen İndüklenmiş Pluripotent Kök Hücreler. Tarih: 25 Eylül 2008 tarihli science-online , doi: 10.1126 / science.1162494 .